
Incidence of Alzheimer’s Disease in Men with Late-Life Hypertension Is Ameliorated by FOXO3 Longevity Genotype
Abstract
Background:
It is well established that mid-life hypertension increases risk of dementia, whereas the association of late-life hypertension with dementia is unclear.
Objective:
To determine whether FOXO3 longevity-associated genotype influences the association between late-life hypertension and incident dementia.
Methods:
Subjects were 2,688 American men of Japanese ancestry (baseline age: 77.0±4.1 years, range 71–93 years) from the Kuakini Honolulu Heart Program. Status was known for FOXO3 rs2802292 genotype, hypertension, and diagnosis of incident dementia to 2012. Association of FOXO3 genotype with late-life hypertension and incident dementia, vascular dementia (VaD) and Alzheimer’s disease (AD) was assessed using Cox proportional hazards models.
Results:
During 21 years of follow-up, 725 men were diagnosed with all-cause dementia, 513 with AD, and 104 with VaD. A multivariable Cox model, adjusting for age, education, APOE ɛ4, and cardiovascular risk factors, showed late-life hypertension increased VaD risk only (HR = 1.71, 95% CI = 1.08–2.71, p = 0.022). We found no significant protective effect of FOXO3 longevity genotype on any type of dementia at the population level. However, in a full Cox model adjusting for age, education, APOE ɛ4, and other cardiovascular risk factors, there was a significant interaction effect of late-life hypertension and FOXO3 longevity genotype on incident AD (β= –0.52, p = 0.0061). In men with FOXO3 rs2802292 longevity genotype (TG/GG), late-life hypertension showed protection against AD (HR = 0.72; 95% CI = 0.55–0.95, p = 0.021). The non-longevity genotype (TT) (HR = 1.16; 95% CI = 0.90–1.51, p = 0.25) had no protective effect.
Conclusion:
This longitudinal study found late-life hypertension was associated with lower incident AD in subjects with FOXO3 genotype.
INTRODUCTION
Hypertension compromises the structural and functional integrity of the cerebral microcirculation, resulting in microvascular rarefaction, endothelial dysfunction, and neurovascular uncoupling, which together impair cerebral blood supply [1]. Cerebral small vessel disease manifests as white matter hyperintensities, lacunar infarcts, microhemorrhages, and accumulation of amyloid-β in perivascular spaces leading to disruption of the blood-brain barrier [2, 3]. The resulting increase in neuroinflammation can lead to cognitive decline [2]. With aging, the deleterious effects of hypertension on the cerebral microvasculature gradually increase, homeostatic functions are increasingly disrupted and resilience to stress declines [2]. As a result, amyloid-related pathologies such as Alzheimer’s disease (AD) ensue [2]. While there is clear evidence that increased diastolic blood pressure (DBP) is associated with biomarkers of AD, the role of systolic blood pressure (SBP) is less clear, although, in contrast to DBP, may protect against AD-associated hippocampal volume reduction [4]. It is therefore considered important to study effects of DBP and SBP on brain aging and disease separately [4].
While most [5–11], but not all [12], studies have found that mid-life hypertension increases the risk of dementia and AD, mixed results have been obtained for the association of late-life hypertension with AD. These include an increased risk [13, 14], decreased risk [15–17] (especially in centenarians and near-centenarians [15, 18], or no association [19–23]. It has, moreover, been found that very high SBP is associated with AD, but high DBP is not [24]. In contrast, extremely low DBP was associated with AD in the latter study. In a long-term US follow-up study, sustained hypertension in midlife to late life, as well as midlife hypertension and late-life hypotension, were each associated with risk of AD [25]. A meta-analysis found elevated DBP variability, elevated SBP variability, and orthostatic hypotension were associated with increased risk of AD in the elderly [9]. The apparent inconsistencies in findings for late-life hypertension on AD risk have been puzzling researchers and the public health sector for decades. We therefore wondered whether another factor, such as genetics, might be modulating the impact of late-life hypertension on AD risk, thereby potentially explaining these conflicting observations.
The forkhead/winged helix box O member 3 transcription factor FOXO3 controls multiple pathways involved in healthy aging and longevity [26]. FOXO3 has a protective effect on vascular aging by regulation of pathways that suppress vascular smooth muscle cell proliferation and neointimal hyperplasia [27]. Activation of FOXO3 transcription in human embryonic stem cells resulted in reinforcement of human vascular cell homeostasis, delayed aging, and increased resistance to oxidative injury [28]. As seen in loss-of-function studies, FOXO3 helps to maintain homeostasis of a diverse array of vascular cell types [29, 30]. Thus, FOXO3 may protect against fibrinoid necrosis in cerebral arterioles of patients with chronic hypertension.
Minor alleles of multiple single nucleotide polymorphisms (SNPs) located in the FOXO3 gene (FOXO3), particularly the G-allele of SNP rs2802292, have been strongly associated with human longevity in multiple studies [26, 31, 32]. In the Kuakini Honolulu Heart Program (Kuakini HHP) cohort, the presence of the longevity associated FOXO3 allele was associated with increased likelihood of living to almost 100 years [33]. It has been postulated that the longevity-associated FOXO3 G-allele may confer “resilience” by mitigating the adverse effects of chronic cardiometabolic stress on intracellular processes, thereby reducing the risk of life-threatening cardiovascular events [26, 33]. In support, we found that FOXO3 longevity genotype mitigates the increased risk of mortality posed by having a cardiometabolic disease [34].
The aim of the present study was to determine whether late-life hypertension affects the risk of developing incident all-cause dementia, AD, and vascular dementia (VaD) differently in men with contrasting FOXO3 genotypes.
METHODS
Design
The Kuakini HHP is a population-based longitudinal study that started on the island of Oahu, Hawaii in 1965 with 8,006 resident middle-aged American men of Japanese ancestry followed for the development of coronary heart disease (CHD) and stroke [35–37].
Participants
Participants were identified using World War II Selective Service Registration files. They were aged 45 to 68 years at the baseline examination that took place between 1965 and 1968. The Kuakini Honolulu-Asia Aging Study (Kuakini HAAS) commenced with Kuakini HHP examination 4 (1991–1993) when participants were 71–93 years old. The primary aim of the Kuakini HAAS was to study cognitive function and dementia. At examination 4, the cognitive abilities screening instrument (CASI) was given to 3,734 Kuakini HHP survivors. Procedures performed were in accord with institutional guidelines and were approved by the Institutional Review Board of the Kuakini Medical Center. Written informed consent was obtained at all examination cycles.
Data collection
The sample used for analysis included 2,688 American men of Japanese ancestry (mean age at examination 4 [baseline], 77.0±4.1 SD years, range 71–93 years) from the Kuakini HHP who had been genotyped for FOXO3 rs2802292 and had available information on incident AD from 1991 to 2012. Cognitive function was assessed by CASI testing. All-cause dementia, AD, and VaD cases were diagnosed from examination 4 (1991–1993) through examination 12 (2011-2012). Details on dementia case finding methods have been published elsewhere [38–40]. All-cause dementia was diagnosed using the Diagnostic and Statistical Manual of Mental Disorders-III-R (DSM-III-R), AD was diagnosed using the National Institute of Neurological Disorders and Stroke and AD and Related Disorders Association (NINDS-ADRDA) criteria, and VaD was diagnosed using the California Alzheimer’s Disease Diagnostic and Treatment Centers (ADDTC) criteria by a team comprising a neurologist and at least one other physician with expertise in dementia diagnosis. Diagnosis was based on history from each participant and a proxy informant, standardized neuropsychological tests, a neurological examination, as well as laboratory tests and brain computed tomography scans when deemed necessary to classify sub-types of dementia. Final diagnosis was assigned by the consensus committee, defining all-cause dementia, as well as probable or possible cases of AD, VaD, or “other” as the primary cause. Secondary causes of dementia were also coded. AD was defined as those who had AD as the sole or primary cause of dementia, and VaD was defined similarly. For incident dementia cases, the time of onset was determined as the mid-point from the date of the examination at which diagnosis was made to the date of the prior examination of the subject. For those who were censored, the date of the last examination the subject participated in was defined as the censored time.
Data on cardiovascular risk factors were obtained at the Kuakini HHP baseline examination (examination 4 in 1991–1993). Hypertension variables at baseline were defined as HTN1: SBP/DBP of ≥140/90 mmHg or the self-reported use of anti-hypertensive medications at Kuakini HHP examination 4; HTN2: SBP/DBP of ≥160/95 mmHg or the self-reported use of anti-hypertensive medications at Kuakini HHP examination 4. Since all participants were aged over 70 years at baseline, those with hypertension had this condition in late life. We defined time of hypertension diagnosis (i.e., hypertension diagnosed at Kuakini HHP examination 3 (1971–1974; age = 60.2±5.5 years), based on SBP/DBP ≥140/90 mmHg or self-reported use of anti-hypertensive medications, as hypertension diagnosed in midlife (T-HTN-ML). Those who were normotensive at examination 3 but at examination 4 had SBP/DBP ≥140/90 mmHg or self-reported the use of anti-hypertensive medications were deemed to have hypertension diagnosed in late-life (T-HTN-LL). Those with SBP/DBP <140/90 at examination 3 and examination 4 were considered to be normotensive (NMT).
Body mass index (BMI) was defined as weight in kilograms divided by height in meters squared. Diabetes was defined by fasting glucose/two-hour oral glucose tolerance test result of ≥126/200 mg/dl or reported use of insulin or oral hypoglycemic medications (modified American Diabetes Association criteria). Smoking was defined as pack-years by self-report. Physical activity index was quantified as metabolic output during a typical 24-hour period by multiplying a weighting factor by the number of hours spent in 5 activity levels (no activity = 1.0, sedentary = 1.1, slight = 1.5, moderate = 2.4, and heavy = 5.0) [41]. Alcohol intake was measured by self-report as ounces per month. Depressive symptoms were measured using the 11-item Center for Epidemiological Studies Depression (CES-D) Scale [37]. Presence of depressive symptoms was defined as a total CES-11 score CESD-11 ≥9.
Genotyping
Seven FOXO3 SNPs in a haplotype block were genotyped using blood samples collected at examination 4. The FOXO3 longevity haplotype is comprised of a minimum of 14 SNPs with putative functional significance that are in a high degree of linkage disequilibrium in the Japanese population [33]. We genotyped 7 SNPs as a surrogate for the complete haplotype. These comprised the following SNPs (minor alleles shown in brackets): rs2802292 (G), rs2253310 (C), rs2802288 (A), rs2764264 (C), rs9398171 (C), rs12212067 (G), and rs3800230 (G). Genotyping of FOXO3 and APOE variants was performed using DNA from buffy coat blood samples that had been frozen at –70°C [42]. Genotyping was performed using TaqMan on an Applied Biosystems QuantStudio 12K Flex system (ThermoFisher Scientific, Waltham, MA, USA).
Statistical analyses
General linear models were used to compare indirect measurements between groups, and logistic models were used to compare the direct measurements at baseline. Mean age-adjusted baseline risk factor levels were compared among subjects with the FOXO3 TT genotype and carriers of the FOXO3 G-allele (TG/GG) of SNP rs2802292. This was done separately among subjects who were normotensive and subjects with late-life hypertension. Cox proportional hazard models were used to assess the association of late-life hypertension and FOXO3 genotype with dementia outcomes. The interaction of BP variables and FOXO3 genotype with dementia outcome was tested in a full Cox model, i.e., a Cox model that included BP variables, FOXO3 genotype and the interaction term of (BP*FOXO3 genotype), adjusting for other covariates. The effects (hazard ratio [HR] and 95% confidence intervals [CI]) of late-life hypertension status on dementia outcomes for different FOXO3 genotypes were estimated using stratified analyses. The Cox proportional hazard assumption was tested for each Cox model. All statistical analyses were performed using the Statistical Analysis System (SAS) version 9.4 (Cary, NC, USA) [43]. Figures were generated using StataCorp 2019 Stata Statistical Software Release 16 (College Station, TX, USA) [44].
RESULTS
In this prospective study of 3,734 participants with cognition assessment at Kuakini HHP examination 4, we excluded 226 prevalent dementia cases at baseline. We also excluded 141 subjects without FOXO3 rs2802292 genotype, and 679 subjects who did not attend any follow-up examinations after Kuakini HHP examination 4, i.e., with unknown incident dementia status. In the end, our analytical sample included 2,688 subjects. In the 21 years of follow-up (mean follow-up time 8.63±5.03 SD years), 725 incident dementia cases were diagnosed, including 513 cases of AD and 104 cases of VaD.
Table 1 compares baseline characteristics of subjects by FOXO3 genotype status and by late-life hypertension status. At baseline, subjects with FOXO3 longevity genotypes were significantly older, had lower stroke and CHD prevalence, and higher diabetes prevalence and fasting plasma glucose levels. Subjects with hypertension were significantly older, had higher BMI, consumed more alcohol, had higher plasma glucose levels, and had higher prevalence of diabetes, stroke, and CHD. Since non-random loss to follow-up by late-life hypertension or FOXO3 genotypes may bias our results, we compared rates of loss to follow-up by late-life hypertension status and FOXO3 genotype status after combining the 2,688 subjects with 679 subjects who were lost to follow-up. No differences were found in rates of loss to follow-up by either hypertension status or FOXO3 genotype status. Therefore, the loss to follow-up in our study did not appear to be influenced by hypertension status or FOXO3 genotype status of the subjects.
Table 1
Characteristics of subjects at baseline, age-adjusted, by hypertension status (HTN2) and FOXO3 rs2802292 genotype
| Variables n | NT 1,237 | HTN2 1,451 | p | rs2802292 TT 1429 | rs2802292 TG/GG 1259 | p |
| FOXO3 TG/GG (%)1 | 46.1 | 47.5 | 0.47 | – | – | – |
| Hypertension (%)1 | – | – | – | 53.3 | 54.7 | 0.47 |
| Age (y±SD)1 | 76.7±3.9 | 77.1±4.1 | 0.031 | 76.7±3.9 | 77.2±4.2 | 0.0024 |
| Education (y) | 10.8±3.1 | 10.7±3.1 | 0.52 | 10.8±3.1 | 10.7±3.2 | 0.41 |
| APOE ɛ4 carriage (%) | 17.8 | 19.2 | 0.37 | 18.5 | 18.7 | 0.89 |
| Cognitive (CASI) score | 86.7±9.1 | 86.5±7.9 | 0.65 | 86.8±8.1 | 86.3±8.9 | 0.17 |
| BMI (kg/m2) | 23.3±3.1 | 24.1±2.9 | <0.0001 | 23.6±3 | 23.8±2.9 | 0.12 |
| Systolic blood pressure (mmHg) | 136.4±13.4 | 160.7±21.4 | <0.0001 | 148.8±21.7 | 150.3±21.9 | 0.070 |
| Diastolic blood pressure (mmHg) | 76.3±8.3 | 84.4±11.1 | <0.0001 | 80.5±10.5 | 80.9±10.9 | 0.27 |
| Smoking (pack-years) | 24.4±33.9 | 26.5±34.1 | 0.13 | 24.9±33.5 | 26.3±34.6 | 0.28 |
| Alcohol consumption (ounces/mo.) | 14.6±31 | 20.9±44.1 | <0.0001 | 17.5±38.1 | 18.5±39.4 | 0.52 |
| Physical activity index | 31.4±4.8 | 31.2±4.6 | 0.41 | 31.3±4.8 | 31.4±4.6 | 0.70 |
| Fasting plasma glucose (mg/dl) | 109.9±25.2 | 115.5±30.5 | <0.0001 | 111.7±26.9 | 114.3±29.8 | 0.022 |
| Diabetes (%) | 22.5 | 33.1 | <0.0001 | 26.6 | 30.1 | 0.045 |
| Stroke (%) | 2.3 | 3.8 | 0.021 | 3.7 | 2.4 | 0.058 |
| CHD (%) | 17.5 | 21.2 | 0.016 | 21 | 17.8 | 0.035 |
| Depressive symptoms | 9.4 | 9.3 | 0.95 | 8.9 | 9.8 | 0.45 |
| Loss to follow-up1,2 (%) | 19.2 | 20.97 | 0.20 | 20.12 | 20.22 | 0.95 |
NT: normotensive; HTN2: SBP/DBP ≥160/95 or on anti-hypertensive medication. 1Not adjusted for age. 2Loss to follow-up by hypertension status and FOXO3 genotypes were based on 2,688 eligible subjects + 679 subjects lost during follow-up.
Table 2
Main effects (HR and 95% confidence intervals) of late-life hypertension status, BP variables, and FOXO3 rs2802292 longevity genotype, on risk of all-cause dementia, Alzheimer’s disease and vascular dementia
| Model A | All-cause dementia | Alzheimer’s disease | Vascular dementia | |||
| Variable | HR (95% CI)1 | p | HR (95% CI) | p | HR (95% CI) | p |
| HTN1 | 1.05 (0.89–1.25) | 0.55 | 0.97 (0.79–1.18) | 0.73 | 1.78 (1.05–2.99) | 0.032 |
| HTN2 | 1.02 (0.88–1.18) | 0.79 | 0.96 (0.81–1.15) | 0.67 | 1.76 (1.16–2.66) | 0.0079 |
| SBP ≥140 | 0.96 (0.83–1.13) | 0.65 | 0.87 (0.73–1.05) | 0.15 | 1.61 (1.03–2.54) | 0.038 |
| SBP ≥160 | 0.96 (0.82–1.13) | 0.63 | 0.84 (0.69–1.02) | 0.072 | 1.88 (1.26–2.79) | 0.0018 |
| DBP ≥90 | 0.96 (0.80–1.16) | 0.68 | 0.89 (0.71–1.11) | 0.30 | 1.50 (0.96–2.32) | 0.074 |
| DBP ≥95 | 0.73 (0.56–0.96) | 0.023 | 0.65 (0.47–0.91) | 0.011 | 1.53 (0.88–2.65) | 0.13 |
| FOXO3_G | 0.88 (0.76–1.03) | 0.10 | 0.87 (0.73–1.04) | 0.12 | 0.77 (0.52–1.14) | 0.19 |
| Haplotype | 0.89 (0.73–1.09) | 0.25 | 0.87 (0.69–1.10) | 0.26 | 0.86 (0.51–1.45) | 0.57 |
| Model B | All-cause dementia | Alzheimer’s disease | Vascular dementia | |||
| Variable | HR (95% CI)* | p | HR (95% CI) | p | HR (95% CI) | p |
| HTN1 | 0.98 (0.82–1.18) | 0.84 | 0.88 (0.71–1.09) | 0.23 | 2.03 (1.11–3.69) | 0.020 |
| HTN2 | 0.93 (0.80–1.09) | 0.39 | 0.88 (0.73–1.06) | 0.19 | 1.71 (1.08–2.71) | 0.022 |
| SBP ≥140 | 0.92 (0.78–1.09) | 0.33 | 0.83 (0.68–1.00) | 0.051 | 1.74 (1.05–2.89) | 0.032 |
| SBP ≥160 | 0.89 (0.75–1.06) | 0.20 | 0.77 (0.63–0.95) | 0.015 | 1.80 (1.17–2.78) | 0.0076 |
| DBP ≥90 | 0.92 (0.75–1.12) | 0.39 | 0.84 (0.67–1.07) | 0.16 | 1.62 (1.00–2.62) | 0.051 |
| DBP ≥95 | 0.67 (0.50–0.90) | 0.0073 | 0.59 (0.41–0.84) | 0.004 | 1.57 (0.85–2.89) | 0.15 |
| FOXO3_G | 0.88 (0.75–1.03) | 0.099 | 0.87 (0.72–1.05) | 0.14 | 0.75 (0.49–1.16) | 0.19 |
| Haplotype | 0.87 (0.70–1.07) | 0.19 | 0.85 (0.66–1.08) | 0.19 | 0.84 (0.48–1.48) | 0.56 |
Model A: HR (95% CI) were estimated from multivariate Cox models adjusted for baseline variables: age, education (years), CASI score, prevalent stroke, APOE ɛ4, and FOXO3_G (having the G-allele of FOXO3 SNP rs2802292, i.e., genotype TG or GG). Model B: HR (95% CI) were estimated from multivariate Cox models adjusted for baseline variables: age, education (years), APOE ɛ4, CASI score, BMI, plasma glucose, smoking (pack-year), alcohol drinking (ounces/month), physical activity index, and prevalent diabetes, CHD, stroke, and depressive symptoms. HTN1, SBP/DBP ≥140/90 mmHg or the self-reported use of anti-hypertensive medications; HTN2, SBP/DBP ≥160/95 mmHg or the self-reported use of anti-hypertensive medications; FOXO3_G, having the G allele of FOXO3 SNP rs2802292, i.e., genotype TG or GG.
Table 3
Incidence of dementia (per 1,000 person-years) of FOXO3 rs2802292 genotypes stratified by late-life hypertension status
| A | Normotensive | Hypertensive (HTN2) | ||||
| TT | TG/GG | p1 | TT | TG/GG | p1 | |
| Total n values | 667 | 570 | – | 762 | 689 | – |
| All-cause dementia – n | 184 | 153 | – | 198 | 190 | – |
| age-adjusted incidence | 34.4 | 32.3 | 0.60 | 33.8 | 33.1 | 0.24 |
| Alzheimer’s disease – n | 127 | 119 | – | 144 | 123 | – |
| age-adjusted incidence | 23. 5 | 25.1 | 0.58 | 24.3 | 21.3 | 0.036 |
| Vascular dementia – n | 22 | 12 | – | 38 | 32 | – |
| age-adjusted incidence | 4.27 | 2.68 | 0.16 | 6.64 | 5.61 | 0.40 |
| B | Normotensive | Hypertensive (HTN1) | ||||
| TT | TG/GG | p1 | TT | TG/GG | p1 | |
| Total n value | 361 | 313 | – | 1068 | 946 | – |
| All-cause dementia – n | 96 | 82 | – | 286 | 261 | – |
| Age-adjusted incidence | 34.5 | 31.6 | 0.90 | 34.2 | 32.8 | 0.14 |
| Alzheimer’s disease – n | 68 | 65 | – | 203 | 177 | – |
| age-adjusted incidence | 23.5 | 25.0 | 0.40 | 24.2 | 22.2 | 0.071 |
| Vascular dementia – n | 10 | 7 | – | 50 | 37 | – |
| age-adjusted incidence | 4.36 | 2.93 | 0.65 | 6.06 | 4.72 | 0.16 |
1p value for test of incidence rates between FOXO3 genotypes (TT versus TG/GG), estimated using Cox proportional hazard models adjusting for age within each hypertension status. HTN1, SBP/DBP ≥140/90 mmHg or the self-reported use of anti-hypertensive medications; HTN2, SBP/DBP ≥160/95 mmHg or the self-reported use of anti-hypertensive medications.
To illustrate the association of FOXO3 genotype and late-life hypertension with incident dementia outcomes, i.e., all-cause dementia, AD, and VaD, we used the FOXO3 SNP rs2802292 (which is the most replicated SNP used for longevity research) as the representative SNP for FOXO3 genotype in this analysis. Table 2 shows the main effects of FOXO3 G-allele carriers and late-life BP and hypertension on incident all-cause dementia (n = 725), AD (n = 513), and VaD (n = 104) estimated from two multiple Cox regression models, namely: (A) adjusting for baseline variables: age, education (years), CASI score, prevalent stroke, and APOE ɛ4 and (B) adjusting for the baseline variables age, education (years), CASI score, APOE ɛ4, BMI, plasma glucose, smoking (pack-year), alcohol drinking (ounces/month), physical activity index, and prevalent diabetes, CHD, stroke, and depressive symptoms. As expected, most BP variables showed nonsignificant protective effects for all-cause dementia and AD at the population level. However, BP variables were associated with increased risk of VaD. Of note, FOXO3 G-allele carriers showed a protective effect for all dementia outcomes, although this was not statistically significant at the population level.
To study whether the effects of FOXO3 genotype on dementia outcomes differed between HTN1 and HTN2 status, we computed the incidence rates of dementia outcomes for FOXO3 rs2802292 genotypes stratified by late-life hypertension status. The results are shown in Table 3. Among the subjects with HTN2, those with the FOXO3 rs2802292 longevity genotype, TG/GG, had a significantly lower incidence of AD than those with the TT common genotype (p = 0.036). And among subjects with HTN1, those with the FOXO3 rs2802292 longevity genotype, TG/GG, had a lower incidence of AD than those with the TT common genotype (p = 0.071). In contrast, among normotensive subjects there were no genotypic differences in AD incidence.
To examine whether FOXO3 rs2802292 genotype modulates the association of late-life BP with incident all-cause dementia, AD, and VaD, we tested the interaction effect of FOXO3 genotype with the late-life BP variables in the full Cox models (Table 4). This showed significant interactions of FOXO3 rs2802292 G-allele carriage with BP variables only for AD risk. Here, a significant interaction indicated that the late-life BP variables affect AD onset differently between FOXO3 genotypes.
Table 4
Probability (p) values for the interaction term between FOXO3 rs2802292 longevity genotype with late– life BP variables on incident all-cause dementia, Alzheimer’s disease, and vascular dementia
| All– cause dementia | Alzheimer’s disease | Vascular dementia | |
| Interaction term | p † | p | p |
| BP*FOXO3_G | |||
| HTN1*FOXO3_G | 0.10 | 0.051 | 0.25 |
| HTN2*FOXO3_G | 0.070 | 0.0061 | 0.81 |
| SBP140*FOXO3_G | 0.15 | 0.061 | 0.84 |
| SBP160*FOXO3_G | 0.023 | 0.0055 | 0.58 |
| DBP90*FOXO3_G | 0.039 | 0.0021 | 0.31 |
| DBP95*FOXO3_G | 0.65 | 0.51 | 0.57 |
†p value for the interaction term estimated from the full Cox models for late-life BP variable and FOXO3 genotype, adjusting for baseline variables: age, education (years), APOE ɛ4, CASI score, BMI, plasma glucose, smoking (pack– year), alcohol drinking (ounces/month), physical activity index, and prevalent diabetes, CHD, stroke, and depressive symptoms. HTN1, SBP/DBP ≥140/90 mmHg or the self-reported use of anti-hypertensive medications; HTN2: SBP/DBP ≥160/95 mmHg or the self-reported use of anti-hypertensive medications; SBP140, SBP ≥140; SBP160, SBP ≥160; DBP90, DBP ≥90; DBP95, DBP ≥95; *, interaction term; FOXO3_G, rs2802292 (TG/GG).
Fig. 1
HRs and 95% CI of late-life hypertension on incidence of AD by FOXO3 longevity genotype (L) and common genotype (C). The p value (top right) shows statistical significance for interaction of HTN2 with FOXO3 genotype. The analysis compared the hazard ratios (HR) of HTN2 versus NT with incident AD between subjects with FOXO3-C and FOXO3-L. HR and 95% CI were computed using the least square means estimated from the multivariate full Cox model, i.e., including late-life HTN2, FOXO3 genotype, and the interaction term of HTN2 with FOXO3 genotype, adjusting for baseline variables: age, education (years), APOE ɛ4, CASI score, BMI, smoking (pack-year), alcohol drinking (ounces/month), physical activity index, plasma glucose, and prevalent diabetes, CHD, stroke, and depressive symptoms. HTN2 refers to data for subjects with SBP/DBP ≥160/95 mmHg or the self-reported use of anti-hypertensive medications.
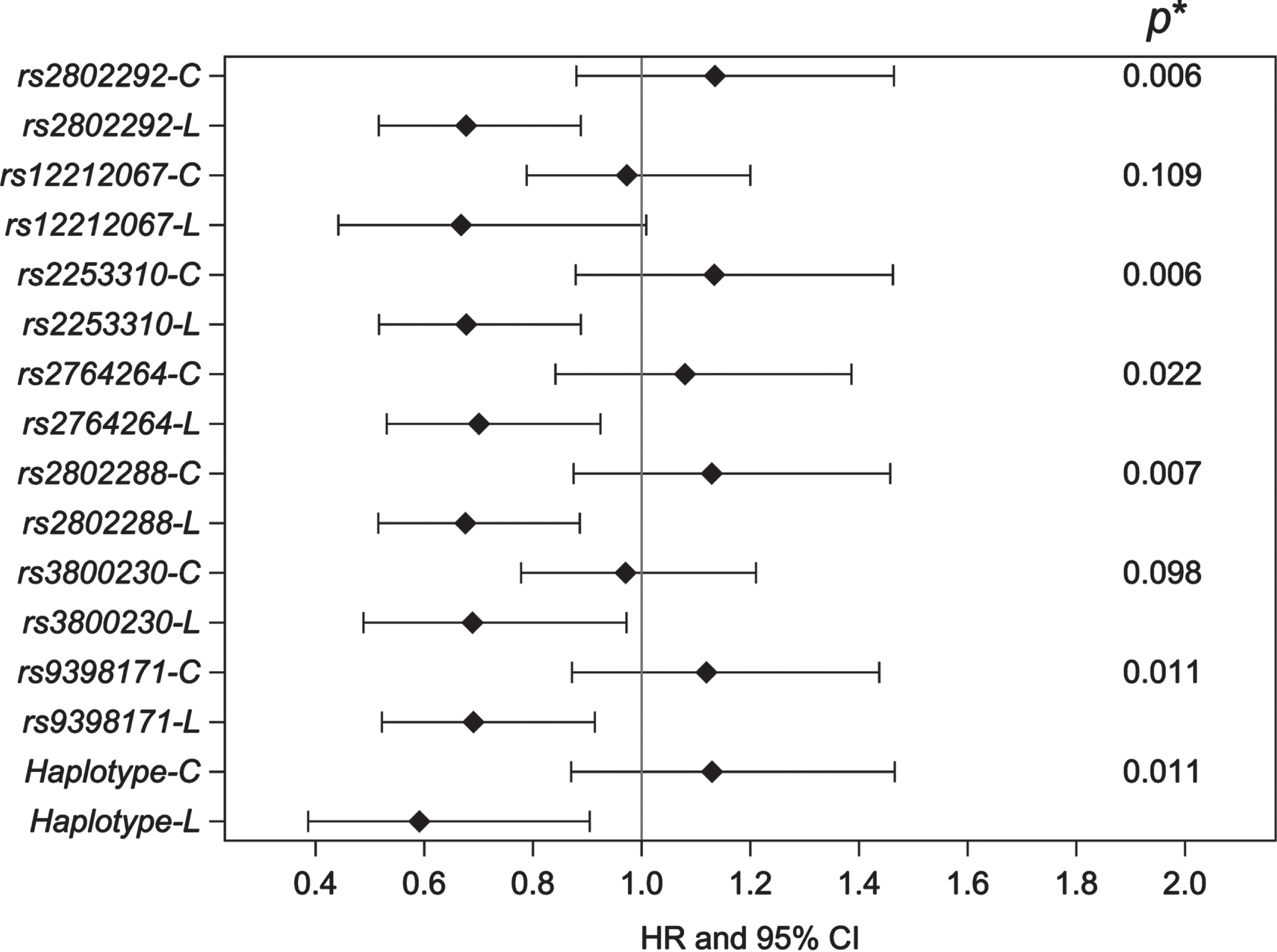
In addition, we examined whether the other six FOXO3 SNPs in LD with each other, as well as the longevity haplotype, reacted to the stress of late-life hypertension so as to affect AD, the major type of dementia. To do this, we tested the interaction effect of HTN2 with each FOXO3 genotype using a multivariate full Cox model adjusting for the baseline variables age, education (years), APOE ɛ4, CASI score, BMI, smoking (pack-year), alcohol drinking (ounces/month), physical activity index, plasma glucose, and prevalent diabetes, CHD, stroke, and depressive symptoms. The p values for interaction tests and the HRs of late-life HTN2 for AD by FOXO3 longevity genotype are presented in Fig. 1. We found a significant interaction effect of late-life HTN2 with the majority of the individual FOXO3 SNPs (5 out of 7), and the FOXO3 longevity haplotype, on AD incidence. For individual FOXO3 longevity genotypes (L), late-life HTN2 was associated with an apparent protective effect against AD risk, with HRs ranging from 0.66 to 0.70. Risk reduction was greatest for the FOXO3 longevity haplotype (HR 0.58; 95% CI 0.38–0.89; p = 0.013). No statistically significant effect was seen for homozygotes of the common allele (C).
Table 5
The effect of late-life BP variables on incident all-cause dementia and Alzheimer’s disease from stratified analyses by FOXO3 rs2802292 genotypes
| All cause dementia | Alzheimer’s disease | ||||
| Genotype | BP variable | HR (95% CI) 1 | p | HR (95% CI) | p |
| TG/GG (n = 1259) | HTN1 | 0.86 (0.66–1.13) | 0.29 | 0.73 (0.53–1.00) | 0.048 |
| HTN2 | 0.85 (0.67–1.08) | 0.18 | 0.72 (0.55–0.95) | 0.021 | |
| SBP ≥140 | 0.84 (0.66–1.08) | 0.18 | 0.71 (0.53–0.95) | 0.019 | |
| SBP ≥160 | 0.78 (0.61–1.00) | 0.051 | 0.61 (0.45–0.83) | 0.0017 | |
| DBP ≥90 | 0.78 (0.58–1.04) | 0.085 | 0.59 (0.41–0.85) | 0.0042 | |
| DBP ≥95 | 0.67 (0.45–1.00) | 0.050 | 0.55 (0.34–0.90) | 0.017 | |
| TT (n = 1429) | HTN1 | 1.15 (0.89–1.48) | 0.28 | 1.09 (0.81–1.46) | 0.56 |
| HTN2 | 1.09 (0.88–1.36) | 0.44 | 1.16 (0.90–1.51) | 0.25 | |
| SBP ≥140 | 1.05 (0.84–1.31) | 0.68 | 1.00 (0.77–1.30) | 0.99 | |
| SBP ≥160 | 1.12 (0.88–1.42) | 0.37 | 1.08 (0.81–1.44) | 0.62 | |
| DBP ≥90 | 1.16 (0.88–1.52) | 0.29 | 1.26 (0.92–1.72) | 0.15 | |
| DBP ≥95 | 0.72 (0.46–1.13) | 0.15 | 0.68 (0.39–1.18) | 0.17 | |
1HR and 95% CI for effect of late-life BP variables on all– cause dementia and Alzheimer’s disease, estimated within each FOXO3 genotype separately from the multivariate Cox models adjusted for baseline variables: age, education (years), APOE ɛ4, CASI score, BMI, plasma glucose, smoking (pack– year), alcohol drinking (ounces/month), physical activity index, and prevalent diabetes, CHD, stroke, and depressive symptoms. HTN1, SBP/DBP ≥140/90 mmHg or the self-reported use of anti-hypertensive medications; HTN2, SBP/DBP ≥160/95 mmHg or the self-reported use of anti-hypertensive medications.
Table 6
Comparisons of effect of untreated hypertension, treatment with anti-hypertensive medication (An), late-life hypertensive (HTN2), and normotensive (NT) status with Alzheimer’s disease incidence, for different FOXO3 rs2802292 genotypes
| Genotype | HTN2 group | Reference group | HR1 (95% CI) | p |
| TG/GG | HTN2 (n = 219) | NT (n = 570) | 0.59 (0.39–0.88) | 0.0099 |
| HTN2 (An) (n = 470) | NT (n = 570) | 0.72 (0.54–0.97) | 0.030 | |
| HTN2 (An) (n = 470) | HTN2 (n = 219) | 1.22 (0.80–1.86) | 0.34 | |
| TT | HTN2 (n = 204) | NT (n = 667) | 1.11 (0.75–1.63) | 0.60 |
| HTN2 (An) (n = 558) | NT (n = 667) | 1.15 (0.87–1.51) | 0.33 | |
| HTN2 (An) (n = 558) | HTN2 (n = 204) | 1.04 (0.70–1.54) | 0.86 |
1HR and (95% CI) was computed using the least square means estimated from the multivariate full Cox models for the HTN2 group and FOXO3 rs2802292 genotype. HRs were compared between HTN2 groups within each genotype (TT, and TG/GG). NT, SBP/DBP <160/95 mmHg; HTN2, untreated hypertension, SBP/DBP ≥160/95 mmHg and not taking anti-hypertensive medication; HTN2 (An), hypertensive subjects who reported taking anti-hypertensive medication.
Table 5 shows the effects of late-life BP variables on incident all-cause dementia and AD, stratified by FOXO3 genotypes. This analysis involved estimating HRs using multivariate Cox models adjusting for age, education, APOE ɛ4 genotype and other cardiovascular risk factors within FOXO3 genotype TT and G-allele carriers separately. For the effects of late-life BP variables on all-cause dementia, only SBP ≥160 mmHg and DBP ≥95 mmHg had a borderline-protective effect in FOXO3 G-allele carriers (HR: 0.78, 95% CI 0.61–1.00; p = 0.051, and HR: 0.67, 95% CI 0.45–1.00; p = 0.050, respectively). In contrast, all BP variables showed statistically significant protective effects against AD in subjects who were FOXO3 G-allele carriers. However, in subjects with the FOXO3 TT genotype all BP variables showed no association with risk of all-cause dementia and AD.
We asked whether the BP measured in mid-life affected this association. Since midlife and late-life BPs were measured approximately 20 years apart, we adjusted for midlife hypertension status as a confounder in the models. Supplementary Table 1 shows that, adjusted for midlife hypertension status, the protective effect of late-life HTN on AD risk still holds for FOXO3 G-allele carriage.
As shown in Supplementary Table 2, after adjustment for major confounding factors (age, education [years], CASI score, prevalent stroke, and APOE ɛ4 genotype), in men with FOXO3 longevity genotype TG/GG, subjects whose hypertension was diagnosed in late life (T-HTN-LL) had significantly reduced risk of developing all-cause dementia (HR = 0.75; p = 0.017), and developing AD (HR = 0.72; p = 0.025), compared to those whose hypertension was diagnosed in midlife (T-HTN-ML).
Since the protective effect of late-life hypertension on risk of dementia may be due to an effect of preclinical dementia that decreased subjects’ blood pressure (a recursive effect) [21], we excluded 53 incident all-cause dementia cases (including 28 AD cases) seen within 1.5 years of follow-up. As shown in Supplementary Table 3, the protective effect of HTN on AD in FOXO3 G-allele carriers remained.
Lowering midlife SBP has been reported to reduce the risk of late-life dementia in the Kuakini HHP-HAAS cohort [45]. To investigate the possible effect of anti-hypertensive medication in late life on AD, we classified the subjects into three late-life BP groups at baseline, namely, normotensive (SBP/DBP <160/95 mmHg), untreated hypertension (SBP/DBP ≥160/95 mmHg, and not taking anti-hypertensive medication), and hypertensive subjects who reported taking anti-hypertensive medication at baseline. Comparisons between the effects of different BP groups on AD by FOXO3 rs2802292 genotype are presented in Table 6. Among subjects who were homozygous for the major allele of FOXO3 rs2802292 (i.e., TT) there were no significant differences among the late-life BP groups and AD incidence (all p > 0.3). In contrast, among subjects having a FOXO3 rs2802292 longevity genotype (TG/GG), there were significant associations in untreated hypertensive and treated hypertensive subjects with protection against AD compared to normotensive subjects (p = 0.0099 and 0.03, respectively). However, the effect of FOXO3 rs2802292 longevity genotype on AD risk reduction showed no significant difference for untreated hypertensive subjects compared with those taking anti-hypertensive medication (p = 0.34).
DISCUSSION
Our study was based on the premise that if the effect of late-life hypertension on AD incidence differed according to FOXO3 genotype, then the overall effect, namely, a weighted average effect of late-life hypertension on AD across different FOXO3 genotypes, would be moving towards the null (i.e., HR = 1). Therefore, the protective effect of late-life hypertension on AD at the population level would be affected by the proportion of FOXO3 G-allele carriers in a population. For the whole Kuakini HHP-HAAS cohort, the HR of the association of late-life hypertension (HTN2) with AD was 0.88 (95% CI: 0.73–1.06; p = 0.19), but in subjects who were carriers of the longevity-associated G-allele, HR was 0.72 (95% CI 0.55–0.95; p = 0.021), and in subjects with the TT genotype, HR was 1.16 (95% CI 0.90–1.51; p = 0.25). Therefore, the protective effect of late-life hypertension on AD in the whole cohort was neutralized by the FOXO3 TT genotype.
We were unable to determine whether the effect of FOXO3 longevity genotype on late-life hypertension-associated AD risk was the result of a single FOXO3 SNP or the combination of genotypes (i.e., the haplotype comprising 7 FOXO3 SNPs). Nevertheless, the association of late-life hypertension-associated AD incidence by FOXO3 longevity haplotype was similar to that from carriage of the representative FOXO3 SNP rs2802292 longevity genotype (TG/GG). It can be expected that at the population level, the protective effect of the FOXO3 longevity haplotype for developing AD will be neutralized (HR = 0.85; p = 0.19) by other FOXO3 haplotypes. Our FOXO3 genotype findings would therefore appear to resolve the conflicting observations concerning the relationship of late-life hypertension with development of incident AD. An implication from our findings is that it is crucial to stratify data by FOXO3 longevity genotype(s) in future research when assessing the association of late-life hypertension with AD risk and in hypertension targeted prevention programs.
What then may be the mechanisms responsible? In non-diabetic patients with uncomplicated hypertension, aortic stiffness is a major determinant of SBP and its variability [46]. Structural and functional changes of the cerebral circulation in patients with hypertension may be caused by an excessive turnover of cerebrovascular fluid, induced by excessive pressure pulsatility, which was termed pulse wave encephalopathy [47]. Although a blood flow regulatory mechanism termed cerebral autoregulation helps to maintain steady cerebral blood flow in response to changes in BP [48], the cerebral circulation is nonetheless torrential and has minimal vascular resistance, making small cerebral arteries particularly susceptible to pressure pulsatility [49], which is amplified by the incompressibility of the skull. Periventricular white matter lesions result, but not deep subcortical lesions. Furthermore, pulsatile cerebrospinal fluid movements may damage the ependymal lining of blood vessels [47], so also contributing to the burden of white matter lesions [50]. A meta-analysis found no overall relationship between pulse pressure and AD [9]. This was explained as stemming from its U-shaped relationship, with both higher and lower tertiles of pulse pressure being associated with increased AD risk [9]. Chronic cerebral hypoperfusion would result in lower BP and appearance of being normotensive.
Increased intraluminal pressure of arterioles in hypertension results in disruption of the blood-brain barrier, increases in reactive oxygen species, weakening of vessel walls, and influx of inflammatory cytokines and neurotoxic blood-derived debris, cells, and microbial pathogens into the central nervous system, resulting in neurodegeneration (see review [51]). Neuroinflammation and activation of microglia play an important role in AD [2, 52]. A circular form of FOXO3 (circFOXO3) has been shown to reduce disruption of the blood-brain barrier following ischemia/reperfusion in the mouse [53]. SNP rs12196996, included in the FOXO3 longevity haplotype, was found to be involved in mRNA processing, including that of circFOXO3, which has been associated with risk of CHD [54]. Changes in white matter (in particular of myelin), oligodendrocytes affected by ischemia, oxidative stress, excitotoxicity, iron overload, amyloid-β toxicity and tauopathy, underlie AD pathology [55].
Although the use of anti-hypertensive medication in midlife has been found to be beneficial for lowering AD risk [56], we found little impact of anti-hypertensive medication use in late life for AD risk reduction compared with subjects whose hypertension was untreated. This may be because in the US only 24% of patients have their hypertension under control [57]. In the SPRINT trial, intensive BP control (to SBP <120 mmHg) had no greater effect on dementia outcome than treatment to SBP <140 mmHg [58]. A meta-analysis of randomized, double-blind, placebo-controlled trials found anti-hypertensive treatment in late-mid and late-life reduced the risk of dementia [59]. Another meta-analysis found a moderate reduction in dementia risk in late-life hypertension patients who used anti-hypertensive medications, irrespective of medication type, and risk reduction increased to 43% for 5 years of use [9]. It has been found that treated hypertensives have an increased amyloid-β burden compared to untreated hypertensives [60]. Hypertension accelerates the deposition of microvascular amyloid-β, leading to neurotoxicity and neuronal death [61]. An earlier study of our cohort found that a similarly high SBP/DBP as in our study (SBP ≥160 mmHg; SBP ≥95 mmHg) was associated with neocortical and hippocampal neurofibrillary tangles, neuritic plaques, and brain atrophy [6]. Other promising therapeutics for AD target mitochondria to reduce reactive oxygen species and stimulate energy production [62, 63]. One is melatonin [64], whose release from glia controls mitochondrial function, reduces inflammation and suppresses hyperphosphorylated tau [65, 66]. Senolytics have shown promise in animal models [67].
Although our previous data has shown that FOXO3 longevity genotype mitigates the increased mortality risk posed by having either late-life hypertension, CHD, stroke, diabetes, or a combination of these at examination 4 [34], the present study found no such genotypic effect. Instead, having a FOXO3 longevity genotype may lower risk of late-life hypertension-associated AD incidence.
In the SPRINT trial, intensive BP control (to SBP <120 mmHg) had no greater effect on dementia outcome than treatment to SBP <140 mmHg [58]. A meta-analysis found a moderate reduction in dementia risk in late-life hypertension patients who used anti-hypertensive medications, irrespective of medication type, and risk reduction increased to 43% for 5 years of use [9]. It has been found that treated hypertensives have an increased amyloid-β burden compared to untreated hypertensives [60]. Hypertension accelerates the deposition of microvascular amyloid-β, leading to neurotoxicity and neuronal death [61]. Our findings should not, however, discourage the use of anti-hypertensive medications, since their use is beneficial for reducing other risks such as from cardiovascular events, mostly myocardial infarction and stroke, in late life.
The strengths of our study were its longitudinal design, long follow-up period, large number of clinical parameters collected at multiple examinations over the course of the study from middle-age to late-life, and the fact that we had data spanning mid-life through late-life. There were also limitations. Although 88% of participants were born in Hawaii, there was a theoretical possibility of confounding of FOXO3 genotype due to geographic origin. All subjects were men, so that we recommend that others perform similar studies in women. Since our subjects were all of Japanese descent, studies in other races should be performed.
Conclusions
The present study found, for the first time, that FOXO3 longevity genotype can moderate the effect of late-life hypertension on AD risk. Thus, without knowing the FOXO3 genotype of patients, one should be cautious about generalizing the effect that late-life hypertension status has on AD incidence in a population. In addition, our study confirmed the risk posed by late-life hypertension on vascular dementia.
ACKNOWLEDGMENTS
The authors thank all study participants and their families for their cooperation, and Ms. Ayako Elliott and Ms. Eva Ardo for assistance with genotyping, and Ms. Hiromi Nakada and Ms. Ka-on Fong for monitoring the vital status of Kuakini HHP participants.
FUNDING
Research reported in this publication was supported by the Kuakini Medical Center, the US National Institutes of Health (contract N01-AG-4-2149, Grants 5 U01 AG019349-05, 5R01AG027060 [Kuakini Hawaii Lifespan Study], 5R01AG038707 [Kuakini Hawaii Healthspan Study], 1P20GM125526-01A1 [Kuakini Center of Biomedical Research Excellence for Clinical and Translational Research on Aging]), and contract N01-HC-05102 from the National Heart, Lung, and Blood Institute.
CONFLICT OF INTEREST
B.J.W. and T.A.D. currently hold US patent 20130295566 entitled “Method of using FOXO3A polymorphisms and haplotypes to predict and promote healthy aging and longevity.”
DATA AVAILABILITY
The data presented are available from author Kamal Masaki upon reasonable request.
SUPPLEMENTARY MATERIAL
[1] The supplementary material is available in the electronic version of this article: https://dx.doi.org/10.3233/JAD-230350.
REFERENCES
[1] | Santisteban MM , Iadecola C , Carnevale D ((2023) ) Hypertension, neurovascular dysfunction, and cognitive impairment. Hypertension 80: , 22–34. |
[2] | Ungvari Z , Toth P , Tarantini S , Prodan CI , Sorond F , Merkely B , Csiszar A ((2021) ) Hypertension-induced cognitive impairment: From pathophysiology to public health. Nat Rev Nephrol 17: , 639–654. |
[3] | Montagne A , Zhao Z , Zlokovic BV ((2017) ) Alzheimer’s disease: A matter of blood-brain barrier dysfunction? J Exp Med 214: , 3151–3169. |
[4] | McNeil CJ , Myint PK , Sandu AL , Potter JF , Staff R , Whalley LJ , Murray AD ((2018) ) Increased diastolic blood pressure is associated with MRI biomarkers of dementia-related brain pathology in normative ageing. Age Ageing 47: , 95–100. |
[5] | Launer LJ , Ross GW , Petrovitch H , Masaki K , Foley D , White LR , Havlik RJ ((2000) ) Midlife blood pressure and dementia: The Honolulu-Asia aging study. Neurobiol Aging 21: , 49–55. |
[6] | Petrovitch H , White LR , Izmirilian G , Ross GW , Havlik RJ , Markesbery W , Nelson J , Davis DG , Hardman J , Foley DJ , Launer LJ ((2000) ) Midlife blood pressure and neuritic plaques, neurofibrillary tangles, and brain weight at death: The HAAS. Honolulu-Asia aging Study. Neurobiol Aging 21: , 57–62. |
[7] | Lennon MJ , Koncz R , Sachdev PS ((2021) ) Hypertension and Alzheimer’s disease: Is the picture any clearer? Curr Opin Psychiatry 34: , 142–148. |
[8] | Gottesman RF , Schneider AL , Zhou Y , Coresh J , Green E , Gupta N , Knopman DS , Mintz A , Rahmim A , Sharrett AR , Wagenknecht LE , Wong DF , Mosley TH ((2017) ) Association between midlife vascular risk factors and estimated brain amyloid deposition. JAMA 317: , 1443–1450. |
[9] | Ou YN , Tan CC , Shen XN , Xu W , Hou XH , Dong Q , Tan L , Yu JT ((2020) ) Blood pressure and risks of cognitive impairment and dementia: A systematic review and meta-analysis of 209 prospective studies. Hypertension 76: , 217–225. |
[10] | Malone JE , Elkasaby MI , Lerner AJ ((2022) ) Effects of hypertension on Alzheimer’s disease and related disorders. Curr Hypertens Rep 24: , 615–625. |
[11] | Whitmer RA , Sidney S , Selby J , Johnston SC , Yaffe K ((2005) ) Midlife cardiovascular risk factors and risk of dementia in late life. Neurology 64: , 277–281. |
[12] | Shang X , Hill E , Zhu Z , Liu J , Ge BZ , Wang W , He M ((2021) ) The association of age at diagnosis of hypertension with brain structure and incident dementia in the UK Biobank. Hypertension 78: , 1463–1474. |
[13] | Bermejo-Pareja F , Benito-León J , Louis ED , Trincado R , Carro E , Villarejo A , de la Cámara AG ((2010) ) Risk of incident dementia in drug-untreated arterial hypertension: A population-based study. J Alzheimers Dis 22: , 949–958. |
[14] | Skoog I , Lernfelt B , Landahl S , Palmertz B , Andreasson LA , Nilsson L , Persson G , Odén A , Svanborg A ((1996) ) 15-year longitudinal study of blood pressure and dementia. Lancet 347: , 1141–1145. |
[15] | Corrada MM , Hayden KM , Paganini-Hill A , Bullain SS , DeMoss J , Aguirre C , Brookmeyer R , Kawas CH ((2017) ) Age of onset of hypertension and risk of dementia in the oldest-old: The 90+ study. Alzheimers Dement 13: , 103–110. |
[16] | Forti P , Pisacane N , Rietti E , Lucicesare A , Olivelli V , Mariani E , Mecocci P , Ravaglia G ((2010) ) Metabolic syndrome and risk of dementia in older adults. J Am Geriatr Soc 58: , 487–492. |
[17] | Ogunniyi A , Hall KS , Gureje O , Baiyewu O , Gao S , Unverzagt FW , Smith-Gamble V , Evans RE , Dickens J , Musick BS , Hendrie HC ((2006) ) Risk factors for incident Alzheimer’s disease in African Americans and Yoruba. Metab Brain Dis 21: , 235–240. |
[18] | Leung Y , Barzilai N , Batko-Szwaczka A , Beker N , Boerner K , Brayne C , Brodaty H , Cheung KS , Corrada MM , Crawford JD , Galbussera AA , Gondo Y , Holstege H , Hulsman M , Ishioka YL , Jopp D , Kawas CH , Kaye J , Kochan NA , Lau BH , Lipnicki DM , Lo JW , Lucca U , Makkar SR , Marcon G , Martin P , Meguro K , Milman S , Poon LW , Recchia A , Ribeiro O , Riva E , Rott C , Sikkes SA , Skoog I , Stephan B , Szewieczek J , Teixeira L , Tettamanti M , Wilczyński K , Sachdev P ((2022) ) Cognition, function, and prevalent dementia in centenarians and near-centenarians: An individual participant data (IPD) meta-analysis of 18 studies. Alzheimers Dement 19: , 2265–2275. |
[19] | Raffaitin C , Gin H , Empana JP , Helmer C , Berr C , Tzourio C , Portet F , Dartigues JF , Alpérovitch A , Barberger-Gateau P ((2009) ) Metabolic syndrome and risk for incident Alzheimer’s disease or vascular dementia: The Three-City Study. Diabetes Care 32: , 169–174. |
[20] | Hayden KM , Zandi PP , Lyketsos CG , Khachaturian AS , Bastian LA , Charoonruk G , Tschanz JT , Norton MC , Pieper CF , Munger RG , Breitner JC , Welsh-Bohmer KA ((2006) ) Vascular risk factors for incident Alzheimer disease and vascular dementia: The Cache County study. Alzheimer Dis Assoc Disord 20: , 93–100. |
[21] | Power MC , Weuve J , Gagne JJ , McQueen MB , Viswanathan A , Blacker D ((2011) ) The association between blood pressure and incident Alzheimer disease: A systematic review and meta-analysis. Epidemiology 22: , 646–659. |
[22] | Tyas SL , Manfreda J , Strain LA , Montgomery PR ((2001) ) Risk factors for Alzheimer’s disease: A population-based, longitudinal study in Manitoba, Canada. Int J Epidemiol 30: , 590–597. |
[23] | Ninomiya T, Ohara T, HirakawaY, YoshidaD DoiY, HataJ, KanbaS, IwakiT, KiyoharaY ((2011) ) Midlife and late-life blood pressure and dementia in Japanese elderly: The Hisayama study. Hypertension 58: , 22–28. |
[24] | Qiu C , von Strauss E , Fastbom J , Winblad B , Fratiglioni L ((2003) ) Low blood pressure and risk of dementia in the Kungsholmen project: A 6-year follow-up study. Arch Neurol 60: , 223–228. |
[25] | Walker KA , Sharrett AR , Wu A , Schneider ALC , Albert M , Lutsey PL , Bandeen-Roche K , Coresh J , Gross AL , Windham BG , Knopman DS , Power MC , Rawlings AM , Mosley TH , Gottesman RF ((2019) ) Association of midlife to late-life blood pressure patterns with incident dementia. JAMA 322: , 535–545. |
[26] | Morris BJ , Willcox DC , Donlon TA , Willcox BJ ((2015) ) FOXO3: A major gene for human longevity - a mini-review. Gerontology 61: , 515–525. |
[27] | Abid MR , Yano K , Guo S , Patel VI , Shrikhande G , Spokes KC , Ferran C , Aird WC ((2005) ) Forkhead transcription factors inhibit vascular smooth muscle cell proliferation and neointimal hyperplasia. J Biol Chem 280: , 29864–29873. |
[28] | Yan P , Li Q , Wang L , Lu P , Suzuki K , Liu Z , Lei J , Li W , He X , Wang S , Ding J , Chan P , Zhang W , Song M , Izpisua Belmonte JC , Qu J , Tang F , Liu GH ((2019) ) FOXO3-engineered human ESC-derived vascular cells promote vascular protection and regeneration. Cell Stem Cell 24: , 447-461 e448. |
[29] | Deng L , Huang L , Sun Y , Heath JM , Wu H , Chen Y ((2015) ) Inhibition of FOXO1/3 promotes vascular calcification. Arterioscler Thromb Vasc Biol 35: , 175–183. |
[30] | Zhang H , Zhao Z , Pang X , Yang J , Yu H , Zhang Y , Zhou H , Zhao J ((2017) ) MiR-34a/sirtuin-1/foxo3a is involved in genistein protecting against ox-LDL-induced oxidative damage in HUVECs. Toxicol Lett 277: , 115–122. |
[31] | Willcox BJ , Donlon TA , He Q , Chen R , Grove JS , Yano K , Masaki KH , Willcox DC , Rodriguez B , Curb JD ((2008) ) FOXO3A genotype is strongly associated with human longevity. Proc Natl Acad Sci U S A 105: , 13987–13992. |
[32] | Broer L , Buchman AS , Deelen J , Evans DS , Faul JD , Lunetta KL , Sebastiani P , Smith JA , Smith AV , Tanaka T , Yu L , Arnold AM , Aspelund T , Benjamin EJ , De Jager PL , Eirkisdottir G , Evans DA , Garcia ME , Hofman A , Kaplan RC , Kardia SL , Kiel DP , Oostra BA , Orwoll ES , Parimi N , Psaty BM , Rivadeneira F , Rotter JI , Seshadri S , Singleton A , Tiemeier H , Uitterlinden AG , Zhao W , Bandinelli S , Bennett DA , Ferrucci L , Gudnason V , Harris TB , Karasik D , Launer LJ , Perls TT , Slagboom PE , Tranah GJ , Weir DR , Newman AB , van Duijn CM , Murabito JM ((2015) ) GWAS of longevity in CHARGE consortium confirms APOE and FOXO3 candidacy. J Gerontol A Biol Sci Med Sci 70: , 110–118. |
[33] | Donlon TA , Morris BJ , Chen R , Masaki KH , Allsopp RC , Willcox DC , Elliott A , Willcox BJ ((2017) ) FOXO3 longevity interactome on chromosome 6. Aging Cell 16: , 1016–1025. |
[34] | Chen R , Morris BJ , Donlon TA , Masaki KH , Willcox DC , Davy PMC , Allsopp RC , Willcox BJ ((2020) ) FOXO3 longevity genotype mitigates the increased mortality risk in men with a cardiometabolic disease. Aging (Albany NY) 12: , 23509–23524. |
[35] | Kagan A , Harris BR , Winkelstein W Jr , Johnson KG , Kato H , Syme SL , Rhoads GG , Gay ML , Nichaman MZ , Hamilton HB , Tillotson J ((1974) ) Epidemiologic studies of coronary heart disease and stroke in Japanese men living in Japan, Hawaii and California: Demographic, physical, dietary and biochemical characteristics. J Chronic Dis 27: , 345–364. |
[36] | Worth RM , Kagan A ((1970) ) Ascertainment of men of Japanese ancestry in Hawaii through World War II Selective Service registration. J Chronic Dis 23: , 389–397. |
[37] | Kohout FJ , Berkman LF , Evans DA , Cornoni-Huntley J ((1993) ) Two shorter forms of the CES-D (Center for Epidemiological Studies Depression) depression symptoms index. J Aging Health 5: , 179–193. |
[38] | White L , Petrovitch H , Ross GW , Masaki KH , Abbott RD , Teng EL , Rodriguez BL , Blanchette PL , Havlik RJ , Wergowske G , Chiu D , Foley DJ , Murdaugh C , Curb JD ((1996) ) Prevalence of dementia in older Japanese-American men in Hawaii: The Honolulu-Asia Aging Study. JAMA 276: , 955–960. |
[39] | Havlik RJ , Izmirlian G , Petrovitch H , Ross GW , Masaki K , Curb JD , Saunders AM , Foley DJ , Brock D , Launer LJ , White L ((2000) ) APOE-epsilon4 predicts incident AD in Japanese-American men: The Honolulu-Asia Aging Study. Neurology 54: , 1526–1529. |
[40] | Abbott RD , White LR , Ross GW , Masaki KH , Curb JD , Petrovitch H ((2004) ) Walking and dementia in physically capable elderly men. JAMA 292: , 1447–1453. |
[41] | Abbott RD , Rodriguez BL , Burchfiel CM , Curb JD ((1994) ) Physical activity in older middle-aged men and reduced risk of stroke: The Honolulu Heart Program. Am J Epidemiol 139: , 881–893. |
[42] | Abbott RD , White LR , Ross GW , Petrovitch H , Masaki KH , Snowdon DA , Curb JD ((1998) ) Height as a marker of childhood development and late-life cognitive function: The Honolulu-Asia Aging Study. Pediatrics 102: , 602–609. |
[43] | Statistical Analysis System (SAS) version 9.4. SAS Institute, Cary, NC, USA. https://libguides.library.kent.edu/statconsulting/SAS. Accessed November 24, (2022) . |
[44] | StataCorp L ((2011) ) Stata Statistical Software: Release 12. StataCorp LP, College Station, TX. |
[45] | Launer LJ , Hughes TM , White LR ((2011) ) Microinfarcts, brain atrophy, and cognitive function: The Honolulu Asia Aging Study Autopsy Study. Ann Neurol 70: , 774–780. |
[46] | Schillaci G , Bilo G , Pucci G , Laurent S , Macquin-Mavier I , Boutouyrie P , Battista F , Settimi L , Desamericq G , Dolbeau G , Faini A , Salvi P , Mannarino E , Parati G ((2012) ) Relationship between short-term blood pressure variability and large-artery stiffness in human hypertension: Findings from 2 large databases. Hypertension 60: , 369–377. |
[47] | Bateman GA , Levi CR , Schofield P , Wang Y , Lovett EC ((2008) ) The venous manifestations of pulse wave encephalopathy: Windkessel dysfunction in normal aging and senile dementia. Neuroradiology 50: , 491–497. |
[48] | van Beek AH , Claassen JA , Rikkert MG , Jansen RW ((2008) ) Cerebral autoregulation: An overview of current concepts and methodology with special focus on the elderly. J Cereb Blood Flow Metab 28: , 1071–1085. |
[49] | O’Rourke MF , Safar ME ((2005) ) Relationship between aortic stiffening and microvascular disease in brain and kidney: Cause and logic of therapy. Hypertension 46: , 200–204. |
[50] | Scheltens P , Barkhof F , Leys D , Wolters EC , Ravid R , Kamphorst W ((1995) ) Histopathologic correlates of white matter changes on MRI in Alzheimer’s disease and normal aging. Neurology 45: , 883–888. |
[51] | Sweeney MD , Sagare AP , Zlokovic BV ((2018) ) Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat Rev Neurol 14: , 133–150. |
[52] | Carnevale D , Mascio G , Ajmone-Cat MA , D’Andrea I , Cifelli G , Madonna M , Cocozza G , Frati A , Carullo P , Carnevale L , Alleva E , Branchi I , Lembo G , Minghetti L ((2012) ) Role of neuroinflammation in hypertension-induced brain amyloid pathology. Neurobiol Aging 33: , 205 e219–229. |
[53] | Yang Z , Huang C , Wen X , Liu W , Huang X , Li Y , Zang J , Weng Z , Lu D , Tsang CK , Li K , Xu A ((2022) ) Circular RNA circ-FoxO3 attenuates blood-brain barrier damage by inducing autophagy during ischemia/reperfusion. Mol Ther 30: , 1275–1287. |
[54] | Zhou YL , Wu WP , Cheng J , Liang LL , Cen JM , Chen C , Liu X , Xiong XD ((2020) ) rs12196996, a polymorphism at the gene flanking intron, is associated with circFOXO3 levels and the risk of coronary artery disease. Aging (Albany NY) 12: , 13076–13089. |
[55] | Nasrabady SE , Rizvi B , Goldman JE , Brickman AM ((2018) ) White matter changes in Alzheimer’s disease: A focus on myelin and oligodendrocytes. Acta Neuropathol Commun 6: , 22. |
[56] | Lebouvier T , Chen Y , Duriez P , Pasquier F , Bordet R ((2020) ) Antihypertensive agents in Alzheimer’s disease: Beyond vascular protection. Expert Rev Neurother 20: , 175–187. |
[57] | Centers for Disease Control and Prevention, National Center for Health Statistics ((2021) ) Facts about hypertension. In: Centers for Disease Control andrevention. https://www.cdc.gov/bloodpressure/facts.htm |
[58] | Williamson JD , Pajewski NM , Auchus AP , Bryan RN , Chelune G , Cheung AK , Cleveland ML , Coker LH , Crowe MG , Cushman WC , Cutler JA , Davatzikos C , Desiderio L , Erus G , Fine LJ , Gaussoin SA , Harris D , Hsieh MK , Johnson KC , Kimmel PL , Tamura MK , Launer LJ , Lerner AJ , Lewis CE , Martindale-Adams J , Moy CS , Nasrallah IM , Nichols LO , Oparil S , Ogrocki PK , Rahman M , Rapp SR , Reboussin DM , Rocco MV , Sachs BC , Sink KM , Still CH , Supiano MA , Snyder JK , Wadley VG , Walker J , Weiner DE , Whelton PK , Wilson VM , Woolard N , Wright JT Jr , Wright CB ((2019) ) Effect of intensive vs standard blood pressure control on probable dementia: A randomized clinical trial. JAMA 321: , 553–561. |
[59] | Peters R , Xu Y , Fitzgerald O , Aung HL , Beckett N , Bulpitt C , Chalmers J , Forette F , Gong J , Harris K , Humburg P , Matthews FE , Staessen JA , Thijs L , Tzourio C , Warwick J , Woodward M , Anderson CS ((2022) ) Blood pressure lowering and prevention of dementia: An individual patient data meta-analysis. Eur Heart J 43: , 4980–4990. |
[60] | Ashby EL , Miners JS , Kehoe PG , Love S ((2016) ) Effects of hypertension and anti-hypertensive treatment on amyloid-β (Aβ) plaque load and Aβ-synthesizing and Aβ-degrading enzymes in frontal cortex. J Alzheimers Dis 50: , 1191–1203. |
[61] | Perrotta M , Lembo G , Carnevale D ((2016) ) Hypertension and dementia: Epidemiological and experimental evidence revealing a detrimental relationship. Int J Mol Sci 17: , 347. |
[62] | Tarantini S , Valcarcel-Ares NM , Yabluchanskiy A , Fulop GA , Hertelendy P , Gautam T , Farkas E , Perz A , Rabinovitch PS , Sonntag WE , Csiszar A , Ungvari Z ((2018) ) Treatment with the mitochondrial-targeted antioxidant peptide SS-31 rescues neurovascular coupling responses and cerebrovascular endothelial function and improves cognition in aged mice. Aging Cell 17: , e12731. |
[63] | Whitson JA , Bitto A , Zhang H , Sweetwyne MT , Coig R , Bhayana S , Shankland EG , Wang L , Bammler TK , Mills KF , Imai SI , Conley KE , Marcinek DJ , Rabinovitch PS ((2020) ) SS-31 and NMN: Two paths to improve metabolism and function in aged hearts. Aging Cell 19: , e13213. |
[64] | Sumsuzzman DM , Choi J , Jin Y , Hong Y ((2021) ) Neurocognitive effects of melatonin treatment in healthy adults and individuals with Alzheimer’s disease and insomnia: A systematic review and meta-analysis of randomized controlled trials. Neurosci Biobehav Rev 127: , 459–473. |
[65] | Chen C , Yang C , Wang J , Huang X , Yu H , Li S , Li S , Zhang Z , Liu J , Yang X , Liu GP ((2021) ) Melatonin ameliorates cognitive deficits through improving mitophagy in a mouse model of Alzheimer’s disease. J Pineal Res 71: , e12774. |
[66] | Chen D , Lan G , Li R , Mei Y , Shui X , Gu X , Wang L , Zhang T , Gan CL , Xia Y , Hu L , Tian Y , Zhang M , Lee TH ((2022) ) Melatonin ameliorates tau-related pathology via the miR-504-3p and CDK5 axis in Alzheimer’s disease. Transl Neurodegener 11: , 27. |
[67] | Walaszczyk A , Dookun E , Redgrave R , Tual-Chalot S , Victorelli S , Spyridopoulos I , Owens A , Arthur HM , Passos JF , Richardson GD ((2019) ) Pharmacological clearance of senescent cells improves survival and recovery in aged mice following acute myocardial infarction. Aging Cell 18: , e12945. |


