
Salience Network Functional Connectivity Mediates Association Between Social Engagement and Cognition in Non-Demented Older Adults: Exploratory Investigation
Abstract
Background:
Social engagement has beneficial effects during cognitive aging. Large-scale cognitive brain network functions are implicated in both social behaviors and cognition.
Objective:
We evaluated associations between functional connectivity (FC) of large-scale brain cognitive networks and social engagement, characterized by self-reported social network size and contact frequency. We subsequently tested large-scale brain network FC as a potential mediator of the beneficial relationship between social engagement and cognitive performance.
Methods:
112 older adults (70.7±7.3 years, range 54.6–89.7; 84 women) completed the Lubben Social Network Scale 6 (LSNS-6), National Alzheimer’s Coordinating Center (NACC) Uniform Data Set 3 (UDS-3) cognitive battery, and resting state fMRI. We completed seed-based correlational analysis in the default mode and salience networks. Significant associations between social engagement scores and cognitive performance, as well as between social engagement and FC of brain networks, informed the construction of mediation models.
Results:
Social engagement was significantly associated with executive function and global cognition, with greater social engagement associated with better cognitive performance. Social engagement was significantly associated with salience network FC, with greater social engagement associated with higher connectivity. Salience network FC partially mediated associations between social engagement and both executive function and global cognition.
Conclusions:
Our results suggest that the salience network is a key mediator of the beneficial relationship between social engagement and cognition in older adults.
INTRODUCTION
Social engagement, specifically the size and nature of social networks, is an important determinant of physical and mental health. Older adults are at elevated risk of social isolation and attendant adverse health consequences [1, 2]. In the United States, 24% of community-dwelling adults older than 65 years were socially isolated in 2011 [3], an issue that has received national attention as a public health crisis [4]. Lack of social connection negatively impacts health in a variety of ways, including physical health (cardiovascular disease [5]), emotional health (depression [6]), and cognitive health (cognitive impairment/dementia [7–11]. As the world population ages, dementia incidence is rising and will continue to rise, suggesting a looming health crisis. Identifying modifiable risk factors for dementia, which may be leveraged to alleviate this crisis, is an increasingly critical area of research. Importantly, social engagement can be characterized in a variety of ways, including network size, contact frequency, and emotional closeness with social contacts. Which aspects of social engagement are most relevant for cognitive aging remains under investigation. Larger social networks seem to confer protection against cognitive decline [8, 12, 13], and evidence suggests that increasing social interactions is a promising intervention against cognitive decline [14–16]. However, the biological mechanisms mediating the impacts of social engagement on cognitive functions remains elusive.
The search for neurobiological mediators of this association between social engagement and cognitive function is informed by research on brain regions and networks associated with social functions, and regions and networks associated with cognitive function. Resting state functional magnetic resonance imaging (fMRI) studies have identified several functionally interconnected brain networks that are consistent across individuals [17]. Some of these networks are associated with basic sensory processes (visual, auditory), while others are associated with higher-level cognitive functions [18]. This latter group, large-scale cognitive brain networks, have varying degrees of relevance to social function and individual domains of cognitive performance. Two large-scale networks implicated in social functions are the default mode and salience networks. Functional connectivity (FC) within the default mode network associates with loneliness [19], while regional volumes of salience network nodes associate with social network size [20, 21]. Salience network constituents show structural remodeling following socio-affective training [22]. Appropriate social cognition requires effective shifting from internally-focused to externally-focused attention, and vice versa. The salience network acts as a putative “switch”, detecting salience cues in the environment and facilitating shifts between internally- and externally-focused attention based on those cues [23], suggesting potential relevance of this network to social behavior. The default mode network and salience network are also implicated in cognitive aging. As is true of large-scale networks more broadly, FC within these networks declines in older adulthood [24–26], and associates with cognitive performance in older adults, with greater FC associated with better performance [24, 25, 27]. Furthermore, reduced default mode network functional connectivity is consistently found in participants with probable Alzheimer’s disease [28, 29]. These factors make the default mode and salience networks interesting candidates for mediating the association between social engagement and cognitive function.
While large-scale cognitive network FC can be broadly characterized as a neural correlate of social and cognitive function, its role as mediator of the relationship between social engagement and cognitive performance in older adults is not established. To address this gap, we attempt to answer four questions. First, does social engagement (social network size and contact frequency) associate with cognitive performance, either globally or within specific cognitive domains? Second, does social engagement associate with FC of large-scale cognitive brain networks? Third, does large-scale cognitive network FC mediate the association between social engagement and cognitive functions?
Finally, we perform two sets of exploratory analyses further interrogating our results. To better understand how different aspects of social engagement relate to brain function and cognitive performance, we perform exploratory analyses using sub-scores of our social network scale focused on active social engagement (frequency of contact) and network size for intimate social engagement (emotional close and trusted individuals). To characterize the role of cognitive impairment in these relationships, we examine associations between our variables of interest separately in cognitively unimpaired participants and those with mild cognitive impairment (MCI).
METHODS
Participants
The National Alzheimer’s Coordinating Center (NACC) maintains a repository of the Uniform Data Set (UDS) collected by all the National Institute on Aging (NIA)–funded Alzheimer’s Disease Centers (ADCs) in the United States. There are currently over 30 ADCs. The UDS data includes demographics, medical history, medication use, physical and neurological exam findings, clinical ratings of dementia severity (Clinical Dementia Rating [CDR] Dementia Staging Instrument) [30], and neuropsychological test scores. Systematic guidelines for clinical diagnosis are based on the most up to date published diagnostic research criteria [31–33] and used across the ADCs. Further information on the NACC database may be found at: https://www.alz.washington.edu.html.
We used data from 120 non-demented older adults with MRI recruited from the Ann Arbor and Metro Detroit communities (as of March of 2020), collected by Michigan Alzheimer’s Disease Center (MADC). MADC was funded as one of the NIH-funded ADCs in 2016. All participants used in the current study were free of MRI contraindications and provided informed consent. Data collection and analysis were approved by the University of Michigan IRB (IRBMED # HUM00000382) and Wayne State University IRB (IRB #033719B3X), respectively.
Participants in the current study were selected using the following inclusion criteria: 1) cognitively unimpaired control or a patient with a diagnosis of mild cognitive impairment, 2) over 50 years old, 3) adequate hearing and visual acuity to complete assessments, 4) preference for availability of an informant with regular contact with the participant and was willing to accompany the participant during evaluation and serve as a “study partner or co-participant”. Participants were excluded for any of the following: 1) evidence of a stroke in a clinically relevant area (cerebral cortex or thalamus) or mass lesion on structural brain imaging, if completed, 2) modified Hachinski Ischemic score > 4 [34], 3) clinically significant abnormality on routine screening blood tests (participants may have been enrolled following correction of laboratory abnormality), 4) clinically significant abnormality on routine cerebrospinal fluid laboratory tests, if performed, 5) evidence or history of traumatic brain injury with persistent cognitive deficits, 6) history of central nervous system radiation therapy, 7) history of intellectual disability.
While a cutoff age of 50 years is young to observe cognitive impairment, the Michigan ADRC collects longitudinal data on participants, and using a low cutoff age allows for observation of changes in participants as they transition from midlife adulthood to older adulthood, and potentially from cognitively unimpaired to early onset cognitive impairment. In practical terms, the current study includes data only from a single baseline time point. The youngest participant in our sample is 54 years old, and only five participants were younger than 60 at the time of screening.
Data for eight participants were excluded for excessive motion, characterized as any volume-to-volume motion spike greater than a voxel width (2.4 mm; n = 7) or average framewise displacement greater than 0.5 mm (n = 1). Following these exclusions, a total of 112 participants were included in the current study.
Behavioral data
To characterize social engagement, participants completed the abbreviated Lubben Social Network Scale (LSNS-6) [35], in which participants quantified the size of their social networks which met criteria for contact frequency or intimacy, with separate items asking about relatives and non-relatives. We used the total of ratings from the six items as an overall measure of social engagement. Possible total scores range from 0–30, with higher values representing greater social engagement, and a score lower than 12 indicative of social isolation [35]. To address the distinction between different aspects of social engagement, we created two sub-scores. First, we summed the two items asking about the number social contacts who they see or hear from at least once a month, converted these sums to Z-scores and used this as an active social engagement score. This score more closely reflects the “quantity” of social engagement that occurs at least once a month. Second, we summed the four items asking about number of social contacts with whom they feel close enough to call for help, and with whom they feel at ease enough to discuss private matters, and converted these sums to Z-scores. We used this as an intimate social engagement sub-score. This score more closely reflects the “quality” of social engagement, as the items require a minimum level of intimacy, characterized by emotional closeness and trust. Active and intimate social engagement sub-scores were moderately correlated in this data set (r = 0.581).
To characterize cognitive performance, participants completed the NACC Uniform Data Set 3 (UDS-3) neuropsychological battery [36–38]. Scores for individual tests were transformed to Z-scores using means and standard deviations of cognitively unimpaired UDS participants as of June 2019 (https://www.alz.washington.edu.html/). From these Z-scores, we calculated composite scores for five cognitive domains as described in [37]: executive function, memory, language, attention, and visuospatial. The composite scores were calculated as the average of the Z-scores for their constituent tests. Where noted, the sign of Z-scores was reversed, so for all scores higher values indicate better performance. The executive function composite was composed of Backward Digit Span Test (# of correct trials) and Trail Making Test Part B (# of seconds to complete; reversed). The memory composite was composed of Immediate Craft Story Recall (paraphrase scoring), Delayed Craft Story Recall (paraphrase scoring), and total score for Benson Complex Figure Delayed Recall. The language composite was composed of Category Fluency (animals), Category Fluency (vegetables), Multilingual Naming Test (total score), and Verbal Fluency Phenomic Test (total correct F & L words). The attention composite was composed of Trail Making Test Part A (# of seconds to complete; reversed) and Forward Digit Span Test (# of correct trials). The visuospatial score was composed of the total score for Benson Complex Figure Copy. A global composite score was calculated as the average of the five domain-specific composite scores (see [37] for the details). Higher values for composite scores represented better cognitive performance.
Resting-state functional magnetic resonance imaging
Data acquisition
All scans were conducted at the University of Michigan Functional Magnetic Resonance Imaging Laboratory (Ann Arbor, MI), with a 3-T General Electric MR750 scanner using an eight-channel Head Matrix coil. Acquisition parameters for T1 structural and T2* functional data were in line with the Adolescent Brain Cognitive Development (ABCD) MRI protocol [39]. 3D T1-weighted whole-brain anatomy images were acquired using a magnetization-prepared rapid acquisition gradient-echo sequence: 208 slices, repetition time (TR) = 2500 ms, echo time (TE) = 2.0 ms, flip angle (FA) = 8°, field of view (FOV) = 256 mm, 256×256 voxels, and voxel size = 1 mm×1 mm×1 mm). Functional images were acquired using a T2*-weighted multiband echo-planar imaging sequence: 570 volumes, 60 slices, TR = 800 ms, TE = 30 ms, FA = 52°, voxel size = 2.4 mm×2.4 mm×2.4 mm, multiband acceleration = 6).
Preprocessing
We used FSL_motion_outliers (FSLv6.0.3, FMRIB’s Software Library, https://fsl.fmrib.ox.ac.uk/fsl/; [40]) to determine the maximum and average volume-to-volume framewise displacement for each run. Seven participants with a maximum framewise displacement greater than the voxel width (2.4 mm) and one participant with an average framewise displacement greater than 0.5 mm were excluded from analysis to prevent contamination of group results by motion-induced signal artifacts.
Functional imaging data were motion-corrected with MCFLIRT [41], normalized to the Montreal Neurological Institute (MNI) template, and underwent ICA-AROMA to remove noise-related ICA components from resting-state data [42].
Functional connectivity analysis
Using CONN Toolbox (http://www.nitrc.org/projects/conn, RRID:SCR_009550; [43]), we selected regions of interest for the resting-state networks using the CONN network atlas, derived from Human Connectome Project data. Default mode network included medial prefrontal cortex, posterior cingulate cortex, and left and right lateral parietal cortex. Salience network included dorsal anterior cingulate cortex, left insula and right insula. Sensorimotor network was included as a negative control, as we expected FC in this network not to associate with social engagement. Sensorimotor network included a superior supplementary motor area region, and left and right motor/somatosensory cortex. Data for eight participants was unavailable for the sensorimotor network; analyses for this network therefore included data for 104 participants. We extracted timeseries from each region of interest for each participant and regressed average white matter and cerebrospinal fluid signal from time series. Data underwent bandpass filtering (0.008–0.09) and linear detrending. We calculated r-to-Z-transformed pairwise functional connectivity values for within-network regions of interest, and for each network these pairwise values were averaged to produce a single within-network connectivity value for each participant.
Statistical analysis
Subsequent analyses were performed in SPSS (version 26). We evaluated the association between LSNS-6 total score and UDS-3 global score using bivariate correlation. Upon finding a statistically significant association, we performed post-hoc exploratory analyses to understand if there were specific aspects of social engagement or cognitive performance that were driving the result. These post-hoc analyses used our created active and intimate social engagement sub-scores from the LSNS-6, and the composite scores from the UDS-3. UDS-3 scores which significantly associated with LSNS-6 scores were then included as outcome variables in separate mediation models. Associations between LSNS-6 (total and subscores) and large-scale cognitive network FC were also evaluated using bivariate correlation. Networks whose FC significantly associated with LSNS-6 scores were then included as mediators in subsequent mediation models. We controlled for age and sex. In examining associations to inform our mediation models we set an alpha threshold of 0.05; as our work is exploratory with limited sample size, we did not correct for multiple comparisons. Mediation models were tested using the PROCESS macro in SPSS [44]. Unstandardized indirect effects were computed for 1000 bootstrapped samples, allowing for computation of a 95% confidence interval via indirect effects at the 2.5 and 97.5 percentiles. Mediation was considered significant if the 95% confidence interval did not cross zero. The proportion mediated was calculated as the ratio of the indirect effect to the total effect.
RESULTS
Demographics and neuropsychological scores
For our analysis sample of 112 participants, we calculated sample mean and standard deviation for age in years (70.67±7.32; range: 54.62–89.69), years of education (15.95±2.47), MoCA score (25.42±3.19), and Lubben total score (19.64±4.81). Six participants had a Lubben total score less than 12, which is the suggested clinical cutoff point to indicate social isolation (5.3%). Eighty-four participants were women (75.0%). Thirty-seven were amnestic MCI (33.0%), 13 were non-amnestic MCI (11.6%), and the remaining 62 (55.3%) were those with unimpaired cognition. As expected, participants with MCI performed more poorly than cognitively unimpaired participants on the UDS-3 composite scores for executive function, memory, language, attention, and the global composite (Supplementary Table 1). Forty-two participants were African-American (37.5%) and 69 were European-American (61.6%). Race data was not available for one participant.
Associations between social engagement and cognition
We found a moderate correlation between LSNS-6 total score with global UDS-3 composite score (r = 0.228, p = 0.016), showing that higher social engagement associated with better cognitive performance. Exploratory post-hoc analysis with UDS-3 domain composite scores found a moderate correlation between LSNS-6 total score and executive function UDS-3 composite score (r = 0.282, p = 0.003). Associations with other domains including UDS-3 memory (r = 0.141, p = 0.137), attention (r = 0.185, p = 0.051), language (r = 0.061, p = 0.525), and visuospatial (r = 0.107, p = 0.236) composite scores were not significant (Table 1). We ran additional exploratory post-hoc analysis to examine cognitively unimpaired and MCI participants separately. The association between social engagement and global cognition was not significant in either group, and the association between social engagement and executive function was significant in cognitively unimpaired participants only. However, further analysis using Fisher’s Z showed that the correlation coefficients were not significantly different between diagnostic groups. Further details are reported in the Supplementary Material.
Table 1
Associations of social engagement with cognitive performance and large-scale cognitive network functional connectivity. LSNS-6 total score is significantly associated with executive function, global composite score, and salience network connectivity.
| Correlation between LSNS-6 and: | r | p |
| Cognitive composite scores | ||
| UDS-3 Global | 0.228 | 0.016 |
| UDS-3 Executive Function | 0.282 | 0.003 |
| UDS-3 Memory | 0.141 | 0.137 |
| UDS-3 Attention | 0.185 | 0.051 |
| UDS-3 Language | 0.061 | 0.525 |
| UDS-3 Visuospatial | 0.107 | 0.263 |
| Large-scale brain network connectivity | ||
| Default Mode Network | 0.169 | 0.076 |
| Salience Network | 0.229 | 0.015 |
| Sensorimotor Network (as a negative control) | 0.005 | 0.991 |
Associations between social engagement and network functional connectivity
We found a moderate association between LSNS-6 total score and salience network connectivity (r = 0.229, p = 0.015), such that greater social engagement associated with greater salience network connectivity. Associations with default mode network (r = 0.169, p = 0.076) and our negative control, the sensorimotor network (r = 0.005, p = 0.959) connectivity were not significant (Table 1).
Exploratory post-hoc analysis with LSNS-6 sub-scores found a moderate correlation between the intimate social engagement sub-score and salience network connectivity (r = 0.251, p = 0.007), such that greater intimate social engagement associated with greater salience network connectivity. Association between active engagement score and salience network connectivity was not significant (r = 0.117, p = 0.218). We ran additional exploratory post-hoc analysis examining these associations in cognitively unimpaired and MCI participants separately. Further details are reported in the Supplementary Material.
Mediation models of social engagement/cognitive performance association
We tested three mediation models in which salience network FC mediated the association between social engagement and cognitive performance. The first model used overall social engagement as the predictor variable and global cognition as the outcome variable with the second model using executive function as the outcome variable, based on the results of the exploratory post-hoc test. The results of the exploratory post-hoc test using LSNS-6 sub-scores led us to test a third model, with intimate social engagement was the predictor variable with the global cognition being the outcome variable. Age and sex were included as nuisance covariates.
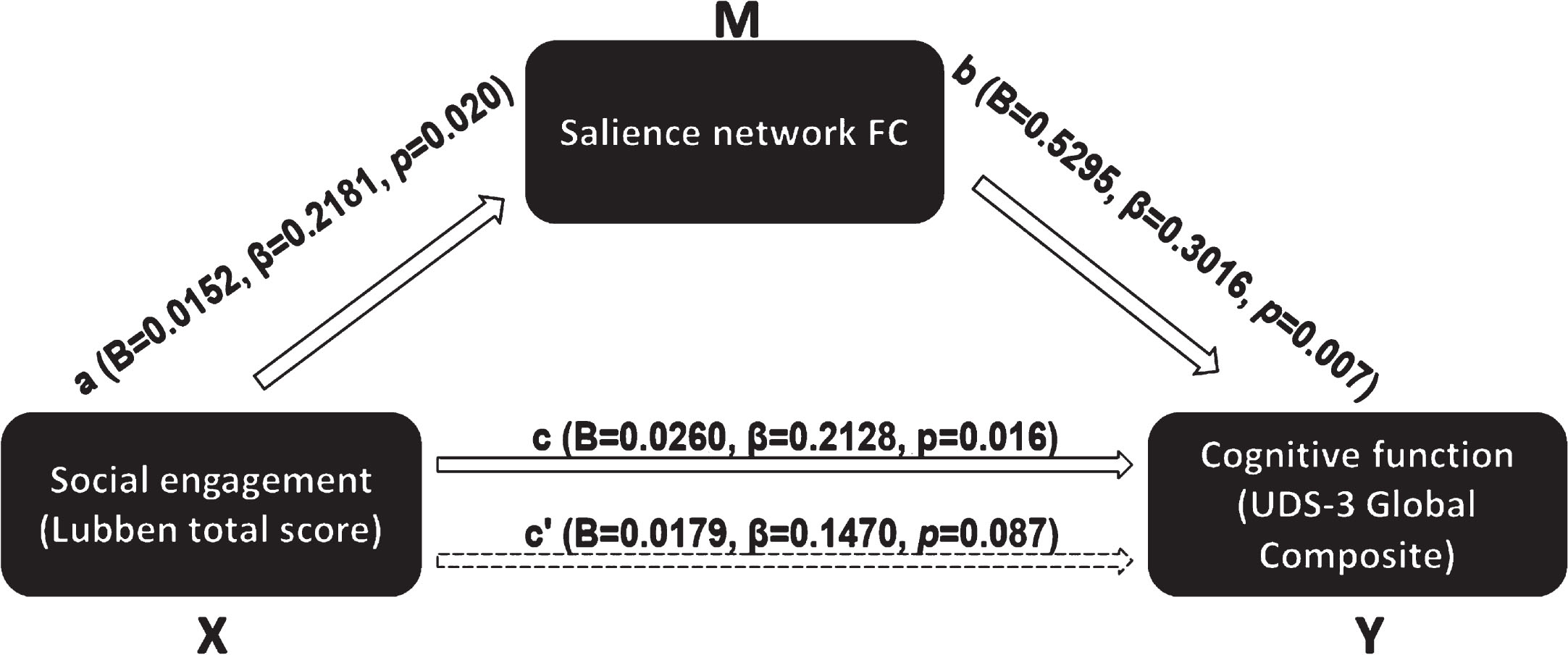
In the first model (Fig. 1), we tested salience network FC as a mediator of the association between LSNS-6 total score and UDS-3 global composite score. The bootstrapped unstandardized indirect effect was 0.0080 (95% CI: 0.0017–0.0169), and salience network functional connectivity mediated 30.8% of the total effect.
Fig. 1
Mediation model of salience network FC mediating the association between overall social engagement and global cognition.
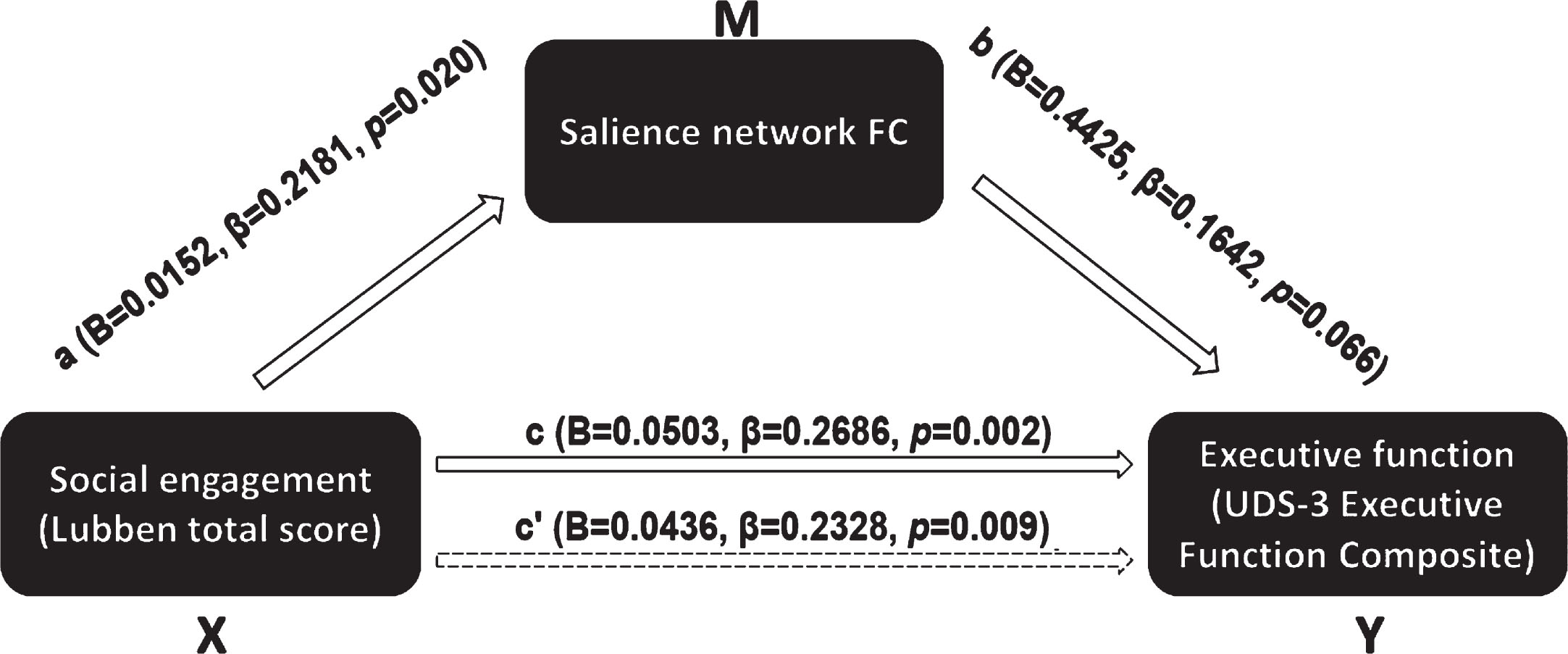
In the second model (Fig. 2), we tested salience network FC as a mediator of the association between LSNS-6 total score and UDS-3 executive function composite score. The unstandardized indirect effect was 0.0067 (95% CI: 0.0000–0.0170), and salience network functional connectivity mediated 13.3% of the total effect.
Fig. 2
Mediation model of salience network FC mediating the association between overall social engagement and executive function.
In the third model, we tested salience network FC as a mediator of the association between LSNS-6 intimate social engagement sub-score and UDS-3 global composite score. The unstandardized indirect effect was 0.0119 (95% CI: 0.0030–0.0241), and salience network functional connectivity mediated 34.7% of the total effect.
We re-ran these three models with additional nuisance covariates, to control for the potential confounding influence of education and race. Educational attainment is related to social engagement, functional connectivity, and cognitive performance. Race, interacting with social conditions not measured here, may also play a role in social engagement and cognitive outcomes. After inclusion of education and race as nuisance covariates, mediation effects for all three models were weakened. Effects for the first and third models remained significant, while the second model no longer showed a significant mediation effect. Additional details are reported in the Supplementary Material.
DISCUSSION
In this study, we evaluated large-scale cognitive network FC as a mediator of associations between social engagement (measured by Lubben’s Social Network Scale-6) and cognitive performance. A global cognitive composite measure was initially used to characterize cognitive performance; exploratory follow-up analyses included five individual cognitive domains: executive function, memory, attention, language, and visuospatial. We tested large-scale cognitive networks as potential mediators: the default mode and salience networks, as well as the sensorimotor network as a negative control. We found a significant association between overall social engagement and the UDS-3 global composite scores. Exploratory post-hoc analyses identified a moderate correlation between overall social engagement and executive function composite score. We also found a significant positive association between overall social engagement and salience network FC. Exploratory post-hoc analyses identified a moderate correlation between intimate social engagement and salience network FC. We found that salience network FC partially mediated associations between overall social engagement and both executive function and global cognition, as well as between intimate social engagement and global cognition. These mediation effects point to the salience network, previously implicated in both social and cognitive functions, as a key mediator of the relationship between social engagement and cognition in older adults.
Our finding of an association between social engagement and cognitive performance in older adults is consistent with prior research. A recent systematic review [45] found that several dimensions of social support (e.g., general social support, emotional support, positive social interaction) are positively associated with global cognition, executive function, and memory [46, 47], though there are dissenting studies [48,49]. While we replicated previously demonstrated positive relationships between social engagement and global and executive function cognitive domains, we did not find a relationship with memory functions. Further study is needed to better understand the factors that impact the association between social engagement and memory performance.
In examining associations between social engagement and large-scale brain network FC, only the association with the salience network was significant. As expected, this association was stronger than the association between social engagement and our negative control, sensorimotor network connectivity, indicating that the relationship with social engagement does not generalize to large-scale brain networks in general. In particular, the network size of intimate social engagement associated with salience network FC. This finding, linking social network size with brain network connectivity is relatively novel: in a recent review of neuroimaging studies of social isolation [50], only one prior study used assessment of social network size and contact frequency [51], and focused on the cerebellum rather than large-scale cognitive networks. Most remaining studies focused on measures of loneliness. Additional work found that social network size was associated with volume and resting-state connectivity of the amygdala [20, 21, 52], a limbic structure frequently included as part of the extended salience network. The specificity of our finding to the salience network may be a function of its role as a hub or “switch” of other large-scale cognitive networks, positioning the network to bridge social function and cognitive performance.
Interestingly, we found that salience network FC was associated with intimate social engagement; that is, the social network size of individuals with whom the person is close and trusts. This finding is exploratory and should be interpreted with caution. As the direction (or presence) of causation is unknown, it may be that a larger intimate social network may contribute to stronger salience network function, or that stronger salience network function facilitates the building of a larger intimate social network. The present study is not equipped to investigate this effect more deeply; nevertheless, our results expand understanding of the understudied neural correlates of social engagement. Notably, previous literature suggests that social engagement with an expansive network of individuals with looser social ties, termed social bridging, is more strongly associated with positive cognitive outcomes than social engagement with a smaller network of individuals with closer ties, termed social bonding [53]. Social bridging is thought to promote cognitive reserve by providing cognitive stimulation through interaction with a diverse set of social contacts, and mitigate the negative impact of reduced amygdala volume [54]. Our results broadly support the idea of increased social engagement promoting cognitive reserve and increased salience network connectivity by providing cognitive stimulation through social interaction. However, our measure of social engagement only allows us to examine this effect in the context of social network size with intimate social contacts. We expect future work to continue to delineate the mechanisms by which social engagement with intimate and peripheral social contacts promote cognitive health in older adults.
Using mediation models, we tested associations between salience network FC and cognitive performance, specifically the global and executive function composite measures. Salience network FC associated with the global cognitive composite score, a finding consistent with recent work demonstrating an association between salience network coherence and a fluid cognition composite score [16]. Hausman et al., describe salience network coherence as associated with several individual subscale scores making up the composite score, including episodic memory, attention, and executive function. In the present study the association between salience network connectivity and executive function was not significant. This discrepancy may be attributable to inclusion of social engagement in our model, which was not included in the Hausman et al.’s model. This further highlights the importance of considering social factors when characterizing the association between FC and cognition.
In exploratory analyses, we examined our questions of interest in our cognitively unimpaired and MCI sub-samples separately. Using Fisher’s Z to assess the significance of the difference between two correlation coefficients between diagnostic groups (i.e. the association between social engagement and brain network connectivity between the two diagnostic groups, and the association between social engagement and cognitive performance between the two diagnostic groups) neither effect was significantly stronger in one group than the other, suggesting that both groups had similar directions and associations in terms of these relationships. We hope future research can provide further clarity on the impact of cognitive impairment on the relationship between social engagement, cognitive performance, and brain network connectivity.
The present study used the Lubben Social Network Scale to measure network size and contact frequency. Another aspect of social function in older adults is captured by perceived social isolation. While conceptually linked, perceived social isolation is distinct from and complementary to measures of social network size and contact frequency. This study, when taken in the context of previous work, suggests that the two may have dissociable neural correlates, with social engagement more strongly linked to the salience network, and perceived social isolation more strongly linked to the default mode network [19]. Further examination of this possible dissociation is warranted and may help shed further light on the neurobiological impact of disrupted social function in older adults.
Several limitations of the approach and sample used in the present study should be noted. First and most significantly, the active and intimate social engagement sub-scores from the LSNS-6 have not been validated and results using these measurements should be treated as exploratory and interpreted with appropriate caution. They are used here to provide exploratory insight into how different aspects of social engagement (size of active social network vs size of intimate social network) relate to brain network functional connectivity and cognitive performance. Furthermore, post-hoc tests were performed when there was a significant association with the LSNS-6 total score. Given that four of the six items were summed for the intimate social engagement score, and only two for the active social engagement score, it is worth noting that the total score is more heavily weighted toward intimate social engagement. It is therefore unsurprising that the post-hoc findings would find significant associations with the intimate social engagement score. Second, we use a cross-sectional approach to examine relationships among our variables of interest, which does not allow us to characterize changes over time. This is of particular importance in the context of cognition, as a major goal of the field is to better understand factors which influence cognitive maintenance and cognitive decline in older age. Building on the present study, future longitudinal work examining the role of social engagement and brain network connectivity in cognitive decline should be valuable. Third, although our participants show a broad range of social engagement in their Lubben scores, our sample as a whole is relatively socially engaged, with only a small subset of participants identified as socially isolated based on Lubben criteria (Lubben 6 item < = 12) [35]. Social isolated subjects might show different patterns of association from those shown here [16]. While our study demonstrates how individual variability in social engagement relates to brain network FC and cognitive performance, examining how these effects hold in a sample with more socially isolated participants would help establish the generalizability of our results. Fourth, while the exclusion criteria did not specifically screen out participants with neuropsychiatric disease such as depression, all participants had low (subclinical) levels of depressive symptoms as measured by the Geriatric Depression Inventory. Late-life depression is often comorbid with cognitive decline [55], accompanied by social isolation [6], and is known to be associated with differences in brain network connectivity [56]. Although we were not able to examine the role of late-life depression in our model due to limited variability, it represents an important factor which may influence how social isolation and brain network FC relate to cognitive aging. Fifth, there are several aspects of functional connectivity analysis for which there are not gold-standard, agreed-upon methodological approaches. One such approach is the parcellation of brain networks. Here, we have opted to use the parcellation included with CONN Toolbox, which focused on a smaller number of core regions for each network and was developed using participants across adult age range rather than older adults specifically. It is unknown the extent to which the selection of different parcellations and numbers of regions may impact the results, or the extent to which results are driven by a single edge. Similarly, we operationalize functional connectivity as the average of the pairwise r-to-Z transformed values between regions of a network. Sixth, we did not perform multiple comparison correction on our exploratory post-hoc analyses. Results from these analyses should be considered exploratory and interpreted with caution. Seventh, our measure of social engagement is limited in several ways. The questions focus only on relatives and friends, and therefore do not capture weaker social ties which may impact cognitive health. For each item, the number of social partners is capped at “9 or more”, meaning variability beyond that level is not captured in our data. The measure focuses on network size based on a minimum frequency of contact or a minimum level of intimacy. However, it does not capture the degree to which the social interactions themselves are “engaging” or satisfying, nor does it capture the types of activities in which social partners engage. Similarly, it does not capture levels of perceived isolation or “loneliness”. Self-report measures of social networks are biased, including by subjective measures like loneliness which are not captured here. We hope future work can further characterize the present findings using more nuanced measurements of social engagement.
In conclusion, our findings demonstrate that associations between social engagement and cognitive performance are mediated by salience network FC. This work supports social engagement, particularly with intimate social contacts, as an important factor influencing cognitive aging and provides evidence for salience network FC as a pathway for this effect. As future studies examine the use of social engagement interventions in delaying or preventing cognitive decline, measurements of brain network FC may help explicate intervention effects and provide insights into individual differences in intervention responses.
AUTHOR CONTRIBUTIONS
Patrick Pruitt (Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Visualization; Writing – original draft; Writing – review & editing); Jessica S. Damoiseaux (Conceptualization; Funding acquisition; Writing – original draft; Writing – review & editing); Benjamin M. Hampstead (Data curation; Funding acquisition; Project administration); Scott J. Peltier (Data curation); Arijit K. Bhaumik (Data curation; Project administration); Roger L. Albin (Writing – review & editing); Hiroko H. Dodge (Conceptualization; Funding acquisition; Supervision; Writing – original draft; Writing – review & editing).
ACKNOWLEDGMENTS
The authors thank Ray Viviano, PhD for support in data processing.
FUNDING
This project was partially supported by the NIH/NIA funded Michigan Alzheimer’s Disease Center (P30AG053760 and P30AG072931), R01AG051628, R01AG056102, and R01AG056712.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available on request from the corresponding author.
SUPPLEMENTARY MATERIAL
[1] The supplementary material is available in the electronic version of this article: https://dx.doi.org/10.3233/ADR-220082.
REFERENCES
[1] | Donovan NJ , Blazer D ((2020) ) Social isolation and loneliness in older adults: Review and commentary of a National Academies Report. Am J Geriatr Psychiatry 28: , 1233–1244. |
[2] | Prohaska T , Burholt V , Burns A , Golden J , Hawkley L , Lawlor B , Leavey G , Lubben J , O’Sullivan R , Perissinotto C , Tilburg T van , Tully M , Victor C , Fried L ((2020) ) Consensus statement: Loneliness in older adults, the 21st century social determinant of health? BMJ Open 10: , e034967. |
[3] | Cudjoe TKM , Roth DL , Szanton SL , Wolff JL , Boyd CM , Thorpe RJ ((2020) ) The Epidemiology of social isolation: National Health and Aging Trends Study. J Gerontol B Psychol Sci Soc Sci 75: , 107–113. |
[4] | Office of the Surgeon General (OSG) ((2023) ) Our Epidemic of Loneliness and Isolation: The U.S. Surgeon General’s Advisory on the Healing Effects of Social Connection and Community, US Department of Health and Human Services, Washington, DC. |
[5] | Valtorta NK , Kanaan M , Gilbody S , Ronzi S , Hanratty B ((2016) ) Loneliness and social isolation as risk factors for coronary heart disease and stroke: Systematic review and meta-analysis of longitudinal observational studies. Heart 102: , 1009–1016. |
[6] | Domènech-Abella J , Mundó J , Haro JM , Rubio-Valera M ((2019) ) Anxiety, depression, loneliness and social network in the elderly: Longitudinal associations from The Irish Longitudinal Study on Ageing (TILDA). J Affect Disord 246: , 82–88. |
[7] | Kuiper JS , Zuidersma M , Zuidema SU , Burgerhof JG , Stolk RP , Oude Voshaar RC , Smidt N ((2016) ) Social relationships and cognitive decline: A systematic review and meta-analysis of longitudinal cohort studies. Int J Epidemiol 45: , 1169–1206. |
[8] | Penninkilampi R , Casey A-N , Singh MF , Brodaty H ((2018) ) The association between social engagement, loneliness, and risk of dementia: A systematic review and meta-analysis. J Alzheimers Dis 66: , 1619–1633. |
[9] | Desai R , John A , Stott J , Charlesworth G ((2020) ) Living alone and risk of dementia: A systematic review and meta-analysis. Ageing Res Rev 62: , 101122. |
[10] | Livingston G , Huntley J , Sommerlad A , Ames D , Ballard C , Banerjee S , Brayne C , Burns A , Cohen-Mansfield J , Cooper C , Costafreda SG , Dias A , Fox N , Gitlin LN , Howard R , Kales HC , Kivimäki M , Larson EB , Ogunniyi A , Orgeta V , Ritchie K , Rockwood K , Sampson EL , Samus Q , Schneider LS , Selbæk G , Teri L , Mukadam N ((2020) ) Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 396: , 413–446. |
[11] | Kallianpur KJ , Masaki KH , Chen R , Willcox BJ , Allsopp RC , Davy P , Dodge HH ((2022) ) Weak social networks in late life predict incident Alzheimer’s disease: The Kuakini Honolulu-Asia Aging Study. J Gerontol A Biol Sci Med Sci 78: , 663–672. |
[12] | Sharifian N , Zaheed AB , Morris EP , Sol K , Manly JJ , Schupf N , Mayeux R , Brickman AM , Zahodne LB ((2022) ) Social network characteristics moderate associations between cortical thickness and cognitive functioning in older adults. Alzheimers Dement 18: , 339–347. |
[13] | Zhou S , Song S , Jin Y , Zheng Z-J ((2020) ) Prospective association between social engagement and cognitive impairment among middle-aged and older adults: Evidence from the China Health and Retirement Longitudinal Study. BMJ Open 10: , e040936. |
[14] | Dodge HH , Zhu J , Mattek N , Bowman M , Ybarra O , Wild K , Loewenstein DA , Kaye JA ((2015) ) Web-enabled conversational interactions as a means to improve cognitive functions: Results of a 6-week randomized controlled trial. Alzheimers Dement (N Y) 1: , 1–12. |
[15] | Dodge HH , Ybarra O , Kaye JA ((2014) ) Tools for advancing research into social networks and cognitive function in older adults. Int Psychogeriatr 26: , 533–539. |
[16] | Dodge HH , Yu K , Wu C-Y , Pruitt PJ , Asgari M , Kaye JA , Hampstead BM , Struble L , Potempa K , Lichtenberg P , Croff R , Albin RL , Silbert LC , I-CONECT team ((2023) ) Internet-based conversational engagement randomized controlled clinical trial (I-CONECT) among socially isolated adults 75+years old with normal cognition or MCI: Topline results. Gerontologist, doi: 10.1093/geront/gnad147 |
[17] | Damoiseaux JS , Rombouts SARB , Barkhof F , Scheltens P , Stam CJ , Smith SM , Beckmann CF ((2006) ) Consistent resting-state networks across healthy subjects. Proc Natl Acad Sci U S A 103: , 13848–13853. |
[18] | Bressler SL , Menon V ((2010) ) Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn Sci 14: , 277–290. |
[19] | Spreng RN , Dimas E , Mwilambwe-Tshilobo L , Dagher A , Koellinger P , Nave G , Ong A , Kernbach JM , Wiecki TV , Ge T , Li Y , Holmes AJ , Yeo BTT , Turner GR , Dunbar RIM , Bzdok D ((2020) ) The default network of the human brain is associated with perceived social isolation. Nat Commun 11: , 6393. |
[20] | Kwak S , Joo W-T , Youm Y , Chey J ((2018) ) Social brain volume is associated with in-degree social network size among older adults. Proc Biol Sci 285: , 20172708. |
[21] | Bickart KC , Wright CI , Dautoff RJ , Dickerson BC , Barrett LF ((2011) ) Amygdala volume and social network size in humans. Nat Neurosci 14: , 163–164. |
[22] | Valk SL , Bernhardt BC , Trautwein F-M , Böckler A , Kanske P , Guizard N , Collins DL , Singer T ((2017) ) Structural plasticity of the social brain: Differential change after socio-affective and cognitive mental training. Sci Adv 3: , e1700489. |
[23] | Menon V , Uddin LQ ((2010) ) Saliency, switching, attention and control: A network model of insula function. Brain Struct Funct 214: , 655–667. |
[24] | Andrews-Hanna JR , Snyder AZ , Vincent JL , Lustig C , Head D , Raichle ME , Buckner RL ((2007) ) Disruption of large-scale brain systems in advanced aging. Neuron 56: , 924–935. |
[25] | Onoda K , Ishihara M , Yamaguchi S ((2012) ) Decreased functional connectivity by aging is associated with cognitive decline. J Cogn Neurosci 24: , 2186–2198. |
[26] | Geerligs L , Renken RJ , Saliasi E , Maurits NM , Lorist MM ((2015) ) A brain-wide study of age-related changes in functional connectivity. Cereb Cortex 25: , 1987–1999. |
[27] | Damoiseaux JS , Beckmann CF , Arigita EJS , Barkhof F , Scheltens Ph , Stam CJ , Smith SM , Rombouts SARB ((2008) ) Reduced resting-state brain activity in the “default network” in normal aging. Cereb Cortex 18: , 1856–1864. |
[28] | Binnewijzend MAA , Schoonheim MM , Sanz-Arigita E , Wink AM , van der Flier WM , Tolboom N , Adriaanse SM , Damoiseaux JS , Scheltens P , van Berckel BNM , Barkhof F ((2012) ) Resting-state fMRI changes in Alzheimer’s disease and mild cognitive impairment. Neurobiol Aging 33: , 2018–2028. |
[29] | Balthazar MLF , de Campos BM , Franco AR , Damasceno BP , Cendes F ((2014) ) Whole cortical and default mode network mean functional connectivity as potential biomarkers for mild Alzheimer’s disease. Psychiatry Res 221: , 37–42. |
[30] | Morris JC ((1993) ) The Clinical Dementia Rating (CDR): Current version and scoring rules. Neurology 43: , 2412–2414. |
[31] | Jack CR , Albert MS , Knopman DS , McKhann GM , Sperling RA , Carrillo MC , Thies B , Phelps CH ((2011) ) Introduction to the recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7: , 257–262. |
[32] | Albert MS , DeKosky ST , Dickson D , Dubois B , Feldman HH , Fox NC , Gamst A , Holtzman DM , Jagust WJ , Petersen RC , Snyder PJ , Carrillo MC , Thies B , Phelps CH ((2011) ) The diagnosis of mild cognitive impairment due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7: , 270–279. |
[33] | McKhann GM , Knopman DS , Chertkow H , Hyman BT , Jack CR , Kawas CH , Klunk WE , Koroshetz WJ , Manly JJ , Mayeux R , Mohs RC , Morris JC , Rossor MN , Scheltens P , Carrillo MC , Thies B , Weintraub S , Phelps CH ((2011) ) The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7: , 263–269. |
[34] | Hachinski VC , Iliff LD , Zilhka E , Du Boulay GH , McAllister VL , Marshall J , Russell RW , Symon L ((1975) ) Cerebral blood flow in dementia. Arch Neurol 32: , 632–637. |
[35] | Lubben J , Blozik E , Gillmann G , Iliffe S , von Renteln Kruse W , Beck JC , Stuck AE ((2006) ) Performance of an abbreviated version of the Lubben Social Network Scale among three European community-dwelling older adult populations. Gerontologist 46: , 503–513. |
[36] | Weintraub S , Besser L , Dodge HH , Teylan M , Ferris S , Goldstein FC , Giordani B , Kramer J , Loewenstein D , Marson D , Mungas D , Salmon D , Welsh-Bohmer K , Zhou X-H , Shirk SD , Atri A , Kukull WA , Phelps C , Morris JC ((2018) ) Version 3 of the Alzheimer Disease Centers’ Neuropsychological Test Battery in the Uniform Data Set (UDS). Alzheimer Dis Assoc Disord 32: , 10–17. |
[37] | Dodge HH , Goldstein FC , Wakim NI , Gefen T , Teylan M , Chan KCG , Kukull WA , Barnes LL , Giordani B , Hughes TM , Kramer JH , Loewenstein DA , Marson DC , Mungas DM , Mattek N , Sachs BC , Salmon DP , Willis-Parker M , Welsh-Bohmer KA , Wild KV , Morris JC , Weintraub S , National Alzheimer’s Coordinating Center (NACC) ((2020) ) Differentiating among stages of cognitive impairment in aging: Version 3 of the Uniform Data Set (UDS) neuropsychological test battery and MoCA index scores. Alzheimers Dement (N Y) 6: , e12103. |
[38] | Sachs BC , Steenland K , Zhao L , Hughes TM , Weintraub S , Dodge HH , Barnes LL , Craft S , Parker ML , Goldstein FC ((2020) ) Expanded demographic norms for version 3 of the Alzheimer Disease Centers’ Neuropsychological Test Battery in the Uniform Data Set. Alzheimer Dis Assoc Disord 34: , 191–197. |
[39] | Casey BJ , Cannonier T , Conley MI , Cohen AO , Barch DM , Heitzeg MM , Soules ME , Teslovich T , Dellarco DV , Garavan H , Orr CA , Wager TD , Banich MT , Speer NK , Sutherland MT , Riedel MC , Dick AS , Bjork JM , Thomas KM , Chaarani B , Mejia MH , Hagler DJ , Daniela Cornejo M , Sicat CS , Harms MP , Dosenbach NUF , Rosenberg M , Earl E , Bartsch H , Watts R , Polimeni JR , Kuperman JM , Fair DA , Dale AM , ABCD Imaging Acquisition Workgroup ((2018) ) The Adolescent Brain Cognitive Development (ABCD) study: Imaging acquisition across 21 sites. Dev Cogn Neurosci 32: , 43–54. |
[40] | Jenkinson M , Beckmann CF , Behrens TEJ , Woolrich MW , Smith SM ((2012) ) FSL. Neuroimage 62: , 782–790. |
[41] | Jenkinson M , Bannister P , Brady M , Smith S ((2002) ) Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage 17: , 825–841. |
[42] | Pruim RHR , Mennes M , van Rooij D , Llera A , Buitelaar JK , Beckmann CF ((2015) ) ICA-AROMA: A robust ICA-based strategy for removing motion artifacts from fMRI data. Neuroimage 112: , 267–277. |
[43] | Whitfield-Gabrieli S , Nieto-Castanon A ((2012) ) Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect 2: , 125–141. |
[44] | Hayes A ((2022) ), Introduction to Mediation, Moderation, and Conditional Process Analysis: Third Edition: A Regression-Based Approach. |
[45] | Costa-Cordella S , Arevalo-Romero C , Parada FJ , Rossi A ((2021) ) Social support and cognition: A systematic review. Front Psychol 12: , 637060. |
[46] | Pillemer SC , Holtzer R ((2016) ) The differential relationships of dimensions of perceived social support with cognitive function among older adults. Aging Ment Health 20: , 727–735. |
[47] | Zuelsdorff ML , Koscik RL , Okonkwo OC , Peppard PE , Hermann BP , Sager MA , Johnson SC , Engelman CD ((2019) ) Social support and verbal interaction are differentially associated with cognitive function in midlife and older age. Neuropsychol Dev Cogn B Aging Neuropsychol Cogn 26: , 144–160. |
[48] | Sims RC , Hosey M , Levy S-A , Whitfield KE , Katzel LI , Waldstein SR ((2014) ) Distinct functions of social support and cognitive function among older adults. Exp Aging Res 40: , 40–59. |
[49] | Ayotte BJ , Allaire JC , Whitfield KE ((2013) ) Social support, physical functioning, and cognitive functioning among older African American adults. Neuropsychol Dev Cogn B Aging Neuropsychol Cogn 20: , 494–510. |
[50] | Zovetti N , Rossetti MG , Perlini C , Brambilla P , Bellani M ((2021) ) Neuroimaging studies exploring the neural basis of social isolation. Epidemiol Psychiatr Sci 30: , e29. |
[51] | Wong NML , Shao R , Wu J , Tao J , Chen L , Lee TMC ((2019) ) Cerebellar neural markers of susceptibility to social isolation and positive affective processing. Brain Struct Funct 224: , 3339–3351. |
[52] | Bickart KC , Hollenbeck MC , Barrett LF , Dickerson BC ((2012) ) Intrinsic amygdala-cortical functional connectivity predicts social network size in humans. J Neurosci 32: , 14729–14741. |
[53] | Perry BL , McConnell WR , Peng S , Roth AR , Coleman M , Manchella M , Roessler M , Francis H , Sheean H , Apostolova LA ((2022) ) Social networks and cognitive function: An evaluation of social bridging and bonding mechanisms. Gerontologist 62: , 865–875. |
[54] | Perry BL , Roth AR , Peng S , Risacher SL , Saykin AJ , Apostolova LG ((2022) ) Social networks and cognitive reserve: Network structure moderates the association between amygdalar volume and cognitive outcomes. J Gerontol B Psychol Sci Soc Sci 77: , 1490–1500. |
[55] | Arve S , Tilvis RS , Lehtonen A , Valvanne J , Sairanen S ((1999) ) Coexistence of lowered mood and cognitive impairment of elderly people in five birth cohorts. Aging Clin Exp Res 11: , 90–95. |
[56] | Tadayonnejad R , Ajilore O ((2014) ) Brain network dysfunction in late-life depression: A literature review. J Geriatr Psychiatry Neurol 27: , 5–12. |


