
The therapeutic mechanism of epilepsy seizures in different target areas: Research on a theoretical model
Abstract
BACKGROUND:
The selection of optimal target areas in the surgical treatment of epilepsy is always a difficult problem in medicine.
OBJECTIVE:
We employed a theoretical calculation model to explore the control mechanism of seizures by an external voltage stimulus acting in different nerve nuclei.
METHODS:
Theoretical analysis and numerical simulation were combined.
RESULTS:
The globus pallidus, excitatory pyramidal neurons, striatal D1 neurons, thalamic reticular nucleus and specific relay nuclei were selected, we analyzed that the electrical stimulation has different effects in these target areas.
CONCLUSIONS:
The data selected were reasonable in study, the results may give a theoretical support for similar studies in clinical.
1.Introduction
Deep brain stimulation technology has been proved to be effective in relieving seizures for intractable epilepsy patients [1, 2]. Cortex and thalamus are the main seizure areas of epilepsy [3], and the subthalamic nucleus (STN) is a common stimulus target for treating seizures [4]. Basal ganglia (BG) as an important unit in the brain has closely input and output relationship with cortical and thalamic circuit [5, 6, 7]. Thus, in addition to the STN, other organizations in the BG may also be effective DBS targets in controlling seizures. Vuong and Devergnas reviewed the possible roles of the BG in regulating absence, neocortical seizures and temporal lobe [8]. di Giacopo et al. observed that substantia nigra DBS can inhibit myoclonus for myoclonic seizures [9]. The electrophysiological and lesion results implied that substantia nigra pars reticulata (SNr) anterior can inhibit convulsant in seizures [10]. Guo et al. showed that SNr-DBS can effectively control seizure activities in rats [11]. SNr-DBS can also relieve amygdala-kindled seizures for a long time [12]. Chua et al. found that the quality of life can be improved with DBS of the ventral pallidum (VP) in rat models [13]. Yu et al. found that VP-DBS can reduce seizure activity and behavior in pilocarpine-treated rats [14]. Cheng et al. observed that low-frequency stimulation at the external globus palladium (GPe) can inhibit tonic-colonic generalized seizures and temporal lobe epilepsy [15]. However, the therapeutic mechanism and effect of these targets are different, the choice of the best stimulation target area is an important direction of clinical medicine research in the future, which need the support of theoretical research results. Also, many studies implied that striatum may involve in controlling epileptic seizures [16], however, if the striatum can be taken as an effective target in the DBS, it still needs a lot of theoretical evidences to support.
Abnormal interactions of cortex and thalamus circuits have been considered to be a key factor in inducing epilepsy seizures, recently, some studies have shown that the inhibition effect is also obvious as the external stimulation was exerted in the thalamocortical system directly [17, 18]. Molnar et al. found that thalamic DBS may increase short-interval intracortical inhibition, which is similar to the treatment effect of antiepileptic drugs [19]. Lee et al. shown that using the DBS at the anterior thalamic nuclei can significant relief epileptic symptoms in the patient [20]. Son et al. observed that centromedian thalamic nucleus DBS is effective and safe in controlling refractory seizures [21]. Recently, Elhadd et al. found that generalised epilepsy can be successfully managed by thalamic DBS [22]. Grewal et al. shown that chronic subthreshold cortical stimulation may be effective for focal epilepsy [23]. Young et al. pointed out that the piriform cortex may relieve seizures in focal and generalized epilepsy [24]. Lundström et al. found that chronic subthreshold cortical stimulation can reduce the probability of seizures [25, 26]. However, the control mechanism of seizures is complex, which should be intensively studied in the dynamic model.
Therefore, the mechanism of DBS in epilepsy is still unclear, the choice of the best treatment plan and region is a difficult problem to be solved. In this paper, we explored the treatment principle of absence epilepsy through exerting voltage stimulation in different neural nuclei of brain network. Different mechanisms were derived, which may be the theoretical basis for further research.
2.The calculation model and method
The coupling relation of different populations is presented in Fig. 1. The specific meaning and symbols of this model can be referred from [27, 28, 29, 30]. The red lines and black lines represent excitatory and inhibitory inputs, respectively. The electrical stimulation was exerted in different targets, which were denoted in red.
Figure 1.
Schematic diagram of the calculation model used in this paper, the symbol and biological significance of which can be referred from [27, 28, 29, 30].
![Schematic diagram of the calculation model used in this paper, the symbol and biological significance of which can be referred from [27, 28, 29, 30].](https://ip.ios.semcs.net:443/media/thc/2021/29-S1/thc-29-S1-thc218043/thc-29-thc218043-g001.jpg)
The dynamic equations that described the network structure of the model were derived from previous studies [27, 28, 29, 30, 31, 32]:
The meaning of parameters in the above equations were listed in Table 1 [27, 28, 29]. Through the numerical calculation of the above equations [27, 28], we got the main results of this paper. The default simulation data were derived from previous literature [27, 28, 29, 30, 31, 32], which are in a reasonable range:
Table 1
| Parameter | Meaning | Parameter | Meaning |
|---|---|---|---|
|
| Maximum cortical discharge rate |
| Average discharge threshold (ADT) of the cortical nuclei |
|
| Maximum striatal discharge rate |
| The ADT of the striatum |
|
| Maximum STN discharge rate |
| The ADT of the SRN |
|
| Maximum SNr discharge rate |
| The ADT of the SNr |
|
| Maximum GPe discharge rate |
| The ADT of the TRN |
|
| Maximum TRN discharge rate |
| The ADT of the STN |
|
| Maximum SRN discharge rate |
| The ADT of the GPe |
|
| The delay induced by the receptors |
| The decay time constant of synaptodendritic |
|
| The rising time constant of synaptodendritic |
| The damping rate in cortex |
|
| Change in discharge rate threshold |
| The nonspecific subthalamic input |
|
| The connection weight from the population “b” to the population “a” | ||
Figure 2.
The stimulation V was acted in GPi, EPN, GPe, SRN, TRN and striatum D1 neurons, respectively. The AE may be well relieved by tuning
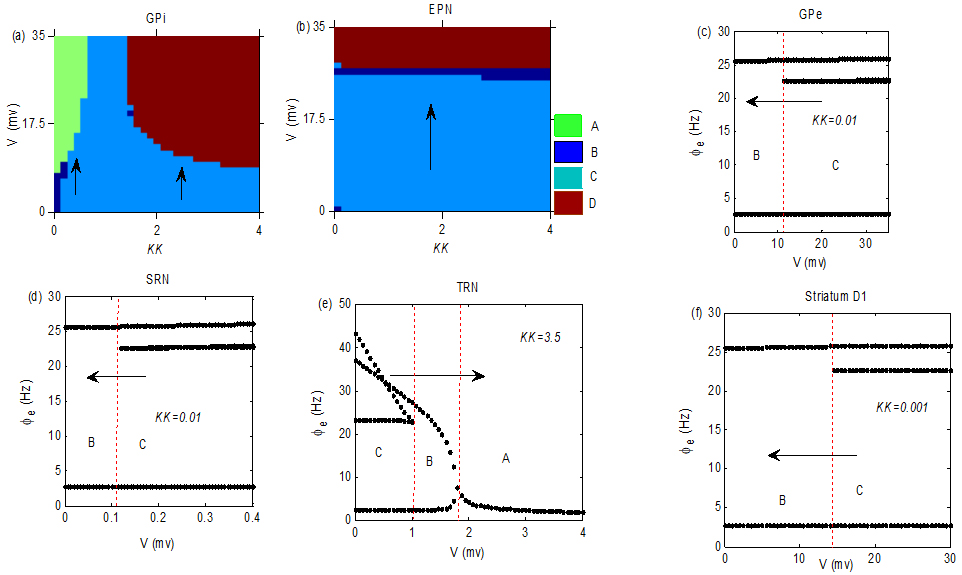
3.Main results
In this section, we explored the control mechanism of seizures by a small voltage V acting in different nuclei. The simulation results were presented in Fig. 2, which is a bifurcation diagram of different states in the EPN. The symbol A represents the low firing state, the B is the simple periodic discharge state, the C is the seizure state and D represents the saturation firing state. The specific meaning and characteristics of these four states can be referred from [27, 28, 29]. KK is a proportional coefficient between “GPi
4.Conclusion
Although there are many reports on the experiment about various potential target areas for the treatment of epilepsy, the theoretical mechanism research based on the model is limited. In this paper, we exerted a stimulation voltage V in the thalamus, GPi, GPe, striatum and EPN respectively to explore the adjustment mechanism of seizures in a dynamic calculation model. We found that positive voltages were suitable for different targets and the stimulus intensity needed to inhibit the seizure was also different. For example, we observed that the intensities needed to inhibit the seizures in SRN and TRN were smaller than that in GPi and EPN. The decrease in V can lead the seizures in Fig. 2c and f to disappear. The parameter values in the model have some influences on the control effect. For example, as shown in Fig. 2a, the excitatory stimulation protocol was effective, but the seizure was transferred to the state A when KK was small, and the seizure was pushed into the state D when KK increased to large enough. As indicated in Fig. 2c and f, the stimulus protocol may be suitable in GPe and striatum D1 when KK was small. The minimum voltage intensity needed to inhibit seizures in SRN and TRN may be relative small as implied in Fig. 2d and e. These different dynamic mechanisms may give theoretical basis for the choice of the optimal scheme and precise treatment in the experiment.
Acknowledgments
This research was supported by the National Science Foundation of China (No. 11602092), the Natural Science Foundation of Hubei Province (No. 2018CFB628), and the China Postdoctoral Science Foundation (No. 2018M632184).
Conflict of interest
None to report.
References
[1] | Zangiabadi N, Ladino LD, Sina F, et al. Deep brain stimulation and drug-resistant epilepsy: a review of the literature. Front Neurol. (2019) ; 10: : 601. |
[2] | Herrman H, Egge A, Konglund AE, et al. Anterior thalamic deep brain stimulation in refractory epilepsy: A randomized, double-blinded study. Acta Neurol. Scand. (2019) ; 139: (3): 294-304. |
[3] | Avoli M, Gloor P. Interaction of cortex and thalamus in spike and wave discharges of feline generalized penicillin epilepsy. Exp. Neurol. (1982) ; 76: (1): 196-217. |
[4] | Charbades S, Kahane P, Minotti L, et al. Deep brain stimulation in epilepsy with particular reference to the subthalamic nucleus. Epileptic disord. (2002) ; 4: : S83-S93. |
[5] | Greene DJ, Marek S, Gordon EM, et al. Integrative and network-specific connectivity of the basal ganglia and thalamus defined in individuals. Neuron. (2020) ; 105: (4): 742-758. |
[6] | Bostan AC, Dum RP, Strick PL. Functional anatomy of basal ganglia circuits with the cerebral cortex and the cerebellum//Current Concepts in Movement Disorder Management. Karger Publishers. (2018) ; 33: : 50-61. |
[7] | Nakajima M, Schmitt LI, Halassa MM. Prefrontal cortex regulates sensory filtering through a basal ganglia-to-thalamus pathway. Neuron. (2019) ; 103: (3): 445-458. |
[8] | Vuong J, Devergnas A. The role of the basal ganglia in the control of seizure. J. Neural Transm. (2018) ; 125: (3): 531-545. |
[9] | di Giacopo A, Baumann CR, Kurthen M, et al. Selective deep brain stimulation in the substantia nigra reduces myoclonus in progressive myoclonic epilepsy: a novel observation and short review of the literature. Epileptic Disord. (2019) ; 21: (3): 283-288. |
[10] | Zhang K, Li Y, Gong H, et al. The role of the substantia nigra pars reticulata anterior in amygdala-kindled seizures. Brain Res. (2019) ; 1715: : 84-93. |
[11] | Guo H, Zhang H, Kuang Y, et al. Electrical stimulation of the substantia nigra pars reticulata (SNr) suppresses chemically induced neocortical seizures in rats. J. Mol. Neurosci. (2014) ; 53: (4): 546-552. |
[12] | Shi LH, Luo F, Woodward D, et al. Deep brain stimulation of the substantia nigra pars reticulata exerts long lasting suppression of amygdala-kindled seizures. Brain Res. (2006) ; 1090: (1): 202-207. |
[13] | Chua FY. The Efficacy of Ventral Pallidum-Deep Brain Stimulation in Rat models of Epilepsy. (2019) . |
[14] | Yu W, Walling I, Smith AB, et al. Deep brain stimulation of the ventral pallidum attenuates epileptiform activity and seizing behavior in pilocarpine-treated rats. Brain Stimul. (2016) ; 9: (2): 285-295. |
[15] | Cheng H, Kuang Y, Liu Y, et al. Low-frequency stimulation of the external globus palladium produces anti-epileptogenic and anti-ictogenic actions in rats. Acta Pharmacol. Sin. (2015) ; 36: (8): 957-965. |
[16] | Rektor I, Kuba R, Brázdil M, et al. Do the basal ganglia inhibit seizure activity in temporal lobe epilepsy? Epilepsy Behav. (2012) ; 25: (1): 56-59. |
[17] | Kokkinos V, Urban A, Sisterson ND, et al. Responsive Neurostimulation of the Thalamus Improves Seizure Control in Idiopathic Generalized Epilepsy: A Case Report. Neurosurg. (2020) . |
[18] | Elder C, Friedman D, Devinsky O, et al. Responsive neurostimulation targeting the anterior nucleus of the thalamus in 3 patients with treatment-resistant multifocal epilepsy. Epilepsia Open. (2019) ; 4: (1): 187-192. |
[19] | Molnar GF, Sailer A, Gunraj CA, et al. Changes in motor cortex excitability with stimulation of anterior thalamus in epilepsy. Neurology. (2006) ; 66: (4): 566-571. |
[20] | Lee CY, Lim SN, Wu T, et al. Successful treatment of refractory status epilepticus using anterior thalamic nuclei deep brain stimulation. World neurosurg. (2017) ; 99: : 14-18. |
[21] | Son B, Shon YM, Choi J, et al. Clinical outcome of patients with deep brain stimulation of the centromedian thalamic nucleus for refractory epilepsy and location of the active contacts. Stereot funct neuros. (2016) ; 94: (3): 187-197. |
[22] | Elhadd KT, Alusi SH, Wieshmann UC, et al. Deep brain thalamic stimulation in the simultaneous treatment of generalised epilepsy and dystonic tremor. Seizure-Eur J Epilep. (2020) ; 75: : 87-88. |
[23] | Grewal SS, Kerezoudis P, Stead M, et al. 216 Chronic Subthreshold Cortical Stimulation for Adult Drug Resistant Focal Epilepsy: Safety, Feasibility, and Technique. Neurosurgery. (2017) ; 64: (CN_suppl_1): 258. |
[24] | Young JC, Vaughan DN, Paolini AG, et al. Electrical stimulation of the piriform cortex for the treatment of epilepsy: a review of the supporting evidence. Epilepsy Behav. (2018) ; 88: : 152-161. |
[25] | Lundström SV, Östman M, Bengtsson-Palme J, et al. Minimal selective concentrations of tetracycline in complex aquatic bacterial biofilms. Sci. Total Environ. (2016) ; 553: : 587-595. |
[26] | Lundstrom B, Van Gompel J, Khadjevand F, et al. Trial stimulation and chronic subthreshold cortical stimulation to treat focal epilepsy. Brain Stimulation: Basic, Translational, and Clinical Research in Neuromodulation, (2019) ; 12: (2): 502. |
[27] | Chen MM, Guo DQ, Wang TB, et al. Bidirectional Control of Absence Seizures by the Basal Ganglia: A Computational Evidence. PLoS Comput. Biol. (2014) ; 10: (3): e1003495. |
[28] | Chen M, Guo D, Li M, et al. Critical roles of the direct GABAergic pallido-cortical pathway in controlling absence seizures. PLoS Comput. Biol. (2015) ; 11: (10): e1004539. |
[29] | Hu B, Guo D, Wang Q. Control of absence seizures induced by the pathways connected to SRN in corticothalamic system. Cogn Neurodynamics. (2015) ; 9: (3): 279-289. |
[30] | Hu B, Wang D, Shi Q. The effect of external voltage stimulation on absence seizures. Technol Health Care. (2020) ; 28: : 245-251. |
[31] | Van Albada SJ, Robinson PA. Mean-field modeling of the basal ganglia thalamocortical system. I Firing rates in healthy and parkinsonian states. J. Theor. Biol. (2009) ; 257: (4): 642-663. |
[32] | van Albada SJ, Gray RT, Drysdale PM, et al. Mean-field modeling of the basal ganglia-thalamocortical system. II: Dynamics of parkinsonian oscillations. J. Theor. Biol. (2009) ; 257: (4): 664-688. |


