
Influence of panoramic cues during prolonged roll-tilt adaptation on the percept of vertical
Abstract
The percept of vertical, which mainly relies on vestibular and visual cues, is known to be affected after sustained whole-body roll tilt, mostly at roll positions adjacent to the position of adaptation. Here we ask whether the viewing of panoramic visual cues during the adaptation further influences the percept of the visual vertical. Participants were rotated in the frontal plane to a 90° clockwise tilt position, which was maintained for 4-minutes. During this period, the subject was either kept in darkness, or viewed panoramic pictures that were either veridical (aligned with gravity) or oriented along the body longitudinal axis. Errors of the subsequent subjective visual vertical (SVV), measured at various tilt angles, showed that the adaptation effect of panoramic cues is local, i.e. for a narrow range of tilts in the direction of the adaptation angle. This distortion was found irrespective of the orientation of the panoramic cues. We conclude that sustained exposure to panoramic and vestibular cues does not adapt the subsequent percept of vertical to the direction of the panoramic cue. Rather, our results suggest that sustained panoramic cues affect the SVV by an indirect effect on head orientation, with a 90° periodicity, that interacts with a vestibular cue to determine the percept of vertical.
1Introduction
Adaptation is a ubiquitous property of all sensory systems, including the visual, vestibular and somatosensory systems. It is well known, for example, that a rocking sensation and postural imbalance is experienced when coming to land after exposure to the sustained motion on a boat [31]. Also, astronauts need a few days to adapt to microgravity in outer space [3, 6, 12, 14, 18] and, back on earth, they initially suffer from spatial disorientation, because the brain’s internal model of the sensory consequences of self-motion is not yet updated to the Earth’s gravity [4, 39].
The otoliths within the vestibular system detect gravity and linear acceleration originating from head motion. Because otolith afferents do not adapt their polarization vectors during prolonged stimulation [11], central pathways must be responsible for behavioral adaptation to microgravity or re-adjustment to normal gravity. Indeed, it has been shown that neurons in the vestibular nuclei adapt their tuning to prolonged head tilts relative to gravity [9, 10]
Early behavioral adaptation studies have shown that vertical estimates drift during prolonged roll-tilt [19, 33]. Furthermore, upon return to an upright position, there is a clear post-tilt bias toward the preceding roll position, which only slowly decays back to zero, likely reflecting a central mechanism [24, 27]. With regards to the generalization of the adaptation, an early study from Higashiyama and Koga (1998) reported that the effects of 10 min tilt-adaptation extend across the whole tilt range [15]. Tarnutzer et al. (2014), however, measuring the SVV in different roll positions after 5 minutes of roll-tilt in one of two adaptation positions (±90°), found that vertical perception adapted only locally, i.e. toward the previously adapted orientation for nearby roll-tilted positions, rather than globally within the entire roll plane [29].
While there is clear effect of vestibular adaptation on the percept of vertical in darkness, in natural conditions there are also visual cues, particularly panoramic cues such as trees, buildings, or the horizon, that are known to affect the percept of vertical [8, 22]. Mismatches between visual and vestibular cues are known to drive central adaptation processes in perception [38] and action [13]. How congruent or incongruent visual and vestibular cues interact in the adaptation of the percept of vertical, i.e. when both cues are presented in combination for a long duration, is not known. In the present study, we therefore test how a visual panoramic cue, polarized to either the direction of gravity or to the body’s long axis, provided continuously during prolonged roll tilt at+90°, affects the subsequent percept of vertical at different head orientations.
From previous work on visuo-vestibular adaptation mechanisms, it is known that if there is a discrepancy between visual and vestibular cues, both cues will adapt in the direction required to reduce the cue conflict [38]. Hence, if the brain adapts the percept of vertical to the polarization direction of the respective visual cues, one expects the gravity-aligned and body-aligned visual cues to induce opposite biases on the error in the percept of vertical compared to prolonged adaptation in the dark.
Alternatively, it has been suggested that the brain interprets a visual panoramic cue as a head orientation cue [1, 20], which explains why such contextual cues typically modulate the SVV with a 90° periodicity [37]. Hence, if a prolonged horizontally or vertically polarized scene causes adaptation of head orientation, such adaptation should also follow the 90° periodicity. As a result, we expect such scenes to induce similar biases in the verticality percept compared to prolonged dark adaptation.
2Materials and methods
2.1Subjects
Nine voluntary participants (4 female, age 30±8 yrs (mean±SD)), provided written informed consent. Each was tested in three experimental sessions of about 90 min each. The protocol was approved by the local ethics committee (Cantonal Ethics committee Zurich, BASEC 2016-00023) and was in accordance with the ethical standards laid down in the 2013 Declaration of Helsinki for research involving human subjects. All participants were free of any known vestibular or neurological disorders and had normal or corrected-to-normal visual acuity. Before the experiment began, subjects were carefully instructed about the task. No feedback about their performance was provided during the experiment. One subject showed poor task performance and was excluded from the study.
2.2Experimental setup
All measurements were performed on a three-axis motor-driven turntable (Acutronic, Jona, Switzerland). The seat of the turntable was adjusted in height to ensure that the center of rotation was aligned with the intersection of the subject’s interaural and naso-occipital axes. Subjects were secured into the seat using a four-point safety belt and a horizontal bar at foot level was used to restrain leg movements. A thermoplastic mask ensured that the subject’s head was securely aligned with the main body axis. Vacuum cushions on both sides of the upper body as well as besides and between the legs and feet minimized head and body motion relative to the turntable.
A turntable-fixed video projector was used to project the panoramic pictures and a laser projected a luminous line (angular subtense 9°) 1.5 m in front of the subject’s eyes, in the center of a dome (3 m diameter) that surrounded the turntable (see [36]). To measure the SVV, the orientation of this line could be adjusted by the subject using a knob, with an accuracy of 0.1°. Subjects viewed the line and the adaptation scene through a circular aperture that limited vision to±36°, preventing any peripheral allocentric cues during visual adaptation. A custom developed Matlab program was used to modify the pictures to avoid distortions when projected onto a spherical surface. A shutter, mounted on the video projector, was closed during the SVV measurement to eliminate any unwanted light.
2.3SVV task
The SVV was tested in darkness at 9 roll tilt angles, ranging from –120 to 120° at 30° intervals. In all conditions, for each participant, the test angle was randomly drawn from this interval, and reached by rotating subjects at a constant acceleration of 10°/s2 followed by a constant deceleration of 10°/s2. After a 5 s waiting period, to let semicircular canal activation dissipate, subjects performed a series of adjustment trials separated by a 1 s pause for a total duration of 1 min. For each trial, the initial orientation of the line was randomly selected and subjects had 5 s to adjust it to their subjective percept of vertical, which they confirmed by a button press. Subjects performed 13±2 (mean±SD) adjustment trials within the 1 minute interval.
Of note, although a forced choice approach may be most preferred for measuring both the perceptual accuracy and precision [5], we used the method of adjustment in this task. The reason is that we needed to limit the time window over which the subject provided responses at each test angle, since adaptation effects may decline. Collecting full psychometric curves for each test angle and each condition (see below) would increase experimental recording time with a factor of 7, which we considered too taxing to our participants. Furthermore, our variable of interest is only the accuracy, not the precision.
2.4Experimental paradigm
The experiment was divided into three experimental sessions of 90 minutes each, yielding > 4.5 hours recording time per participant. Every session started with a baseline condition where SVV was measured without adaptation at all 9 test angles. Subsequently, subjects were tested in one of three conditions, with the order randomized across subjects: 1) adaptation in complete darkness (dark condition); 2) adaptation while viewing a visual panoramic image aligned to gravity (world condition); 3) adaptation while viewing a body-aligned panoramic image (body condition).
2.4.1Baseline condition
Starting from upright, the subject was rotated to the test angle, at which the SVV task was performed in complete darkness. Then the participant was brought to upright for 20 s, and lights were turned on, after which the next orientation was tested, which was repeated up until all angles were assessed.
2.4.2Dark condition
Subjects were rotated in complete darkness from upright to the 90° right ear down tilt position, which was maintained for 240 s (4-minutes). Note, we only tested a single adaption angle, and not the opposite left-ear down angle. Tarnutzer et al. (2014) reported the adaptation effect to be symmetric, and by testing a single adaptation angle we could keep recording time feasible for the participant [29]. Furthermore, we chose a 4-minute adaptation period based on previous observations, including our own, that most of the adaptation effect occur within the first 3–5 min [26, 29, 34]. After this adaptation phase, subjects were rotated to one of the 9 test angles and the SVV task was performed. Subjects then returned to upright for 90 s and lights were turned on to prevent dark adaptation and let the effect of adaptation or hysteresis dissipate (Tarnutzer et al. (2013) have shown that adaptation effects disappear in ∼60s [27]). The whole procedure was repeated nine times, until SVV was measured at all test angles after adaptation. The order of test angles was randomly selected for each participant. Figure 1A shows a representation of the turntable positions during one experimental block.
Fig. 1
A. Tilt angle during one experimental block in the adaptation conditions. Subjects are rotated from upright (0°) to the 90° clockwise tilt position. After an adaptation phase of 240 s, the subject is rotated to a new tilt angle, chosen randomly between ±120°, ±90°, ±60°, ±30° and 0° (i.e. 60° in the figure). After a 5 s delay, the subject performs repetitive SVV adjustments for 60 s, before being brought to upright. Then, 90s later, another trial starts. During the adaptation phase, the subject is either in complete darkness, or presented with panoramic visual cues aligned to the main body axis or aligned to the actual gravitational direction. B. In the World condition a sequence of panoramic images (11 different types), each shown for 10 s, is presented during the adaptation phase, aligned to the gravitational vertical. C. The same sequence of images is shown in the Body condition, aligned to the body longitudinal axis.
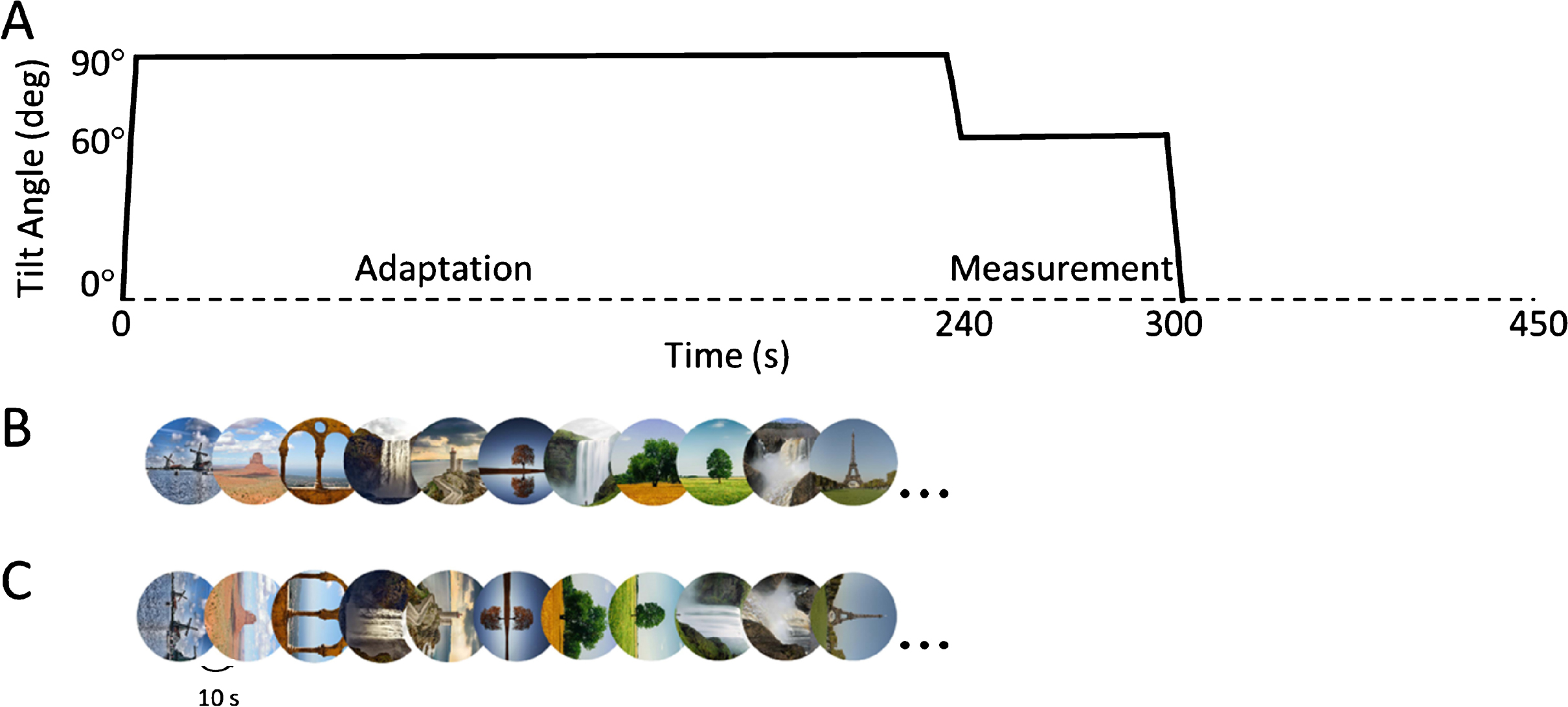
2.4.3World condition
This condition was identical to the dark condition, with the addition that during the adaptation phase a sequence of 24 naturalistic images, each of which randomly chosen from a set of 11 pictures and displayed for 10 s, was shown (Fig. 1B). These images provided clear panoramic cues aligned to the gravitational vertical so that there was a sustained reinforcement of this cue while effects of other, irrelevant visual cues were marginalized across the sequence.
2.4.4Body condition
This condition was identical to the world condition, with the difference that during the adaptation phase subjects viewed the same sequence of naturalistic images but with the panoramic cues aligned to the body’s main axis (Fig. 1C).
2.5Data analysis
Data analyses and statistical tests were conducted using Matlab (MathWorks). For each SVV trial, the adjustment error was quantified as the difference, in degrees, between the final positioning of the line and the true Earth-vertical. There was no systematic temporal drift in the SVV during the one-minute response period (repeated measures ANOVA comparing the first five and last five adjustments for each test angle and experimental condition showed no main effect (F(1,280) = 4.68, p = 0.06) or interaction (F(1, 280) =0.1, p = 0.99)). We therefore considered the SVV measurements as independent adjustments.
For each condition, and for each test angle, accuracy and trial-to-trial variability of the SVV were computed as the median of the adjustment errors and the median absolute deviation (MAD), calculated as the median of the absolute deviations from the median error. Errors deviating > 3 MAD from the median were considered outliers (<3%), and removed from further analysis. Because the baseline conditions did not differ among the three experimental sessions (see Results), errors in the adaptation conditions were compared to the average of the baseline conditions. Furthermore, the SVV errors in the adaptation conditions were analyzed separately for clockwise (0°, 30°, 60°, 90° and 120°) and counterclockwise (0°, –30°, –60°, –90° and –120°) orientations. Because the Jarque-Bera test showed that the errors were normally distributed, we used parametric statistics to address the statistical significance of our effects, including repeated-measures ANOVA’s and t-tests. Holm’s step-down procedure was used to retain the family-wise error rate when multiple comparisons were performed.
3Results
Figure 2A shows the results of a typical subject in the baseline condition, i.e. without effects of static tilt adaptation. For each angle, it shows the sequence of adjustment errors (in gray); the median for each tilt angle is plotted in black. Baseline patterns showing error in the direction opposite to the subject’s tilt angle for small tilt-angles and towards for large tilt angles are seen in all subjects (Fig. 2B), corroborating previous literature [2, 5, 21, 28, 30]. The response curve in the baseline condition was further found to be very stable across sessions: the errors in the three baseline conditions, measured on different days, were not significantly different (F(2,16) = 0.28, p = 0.76, two-way ANOVA with factors angle and session), such that we took the average SVV errors across the three baseline recordings in further analyses.
Fig. 2
SVV errors in the four conditions. Left column: each panel shows the distribution of adjustment errors (gray dots) of a single subject at the various tilt angles; the black dot represents their median. Right column: each panel shows the error curves of single subjects (in gray), as well as their mean (in black).
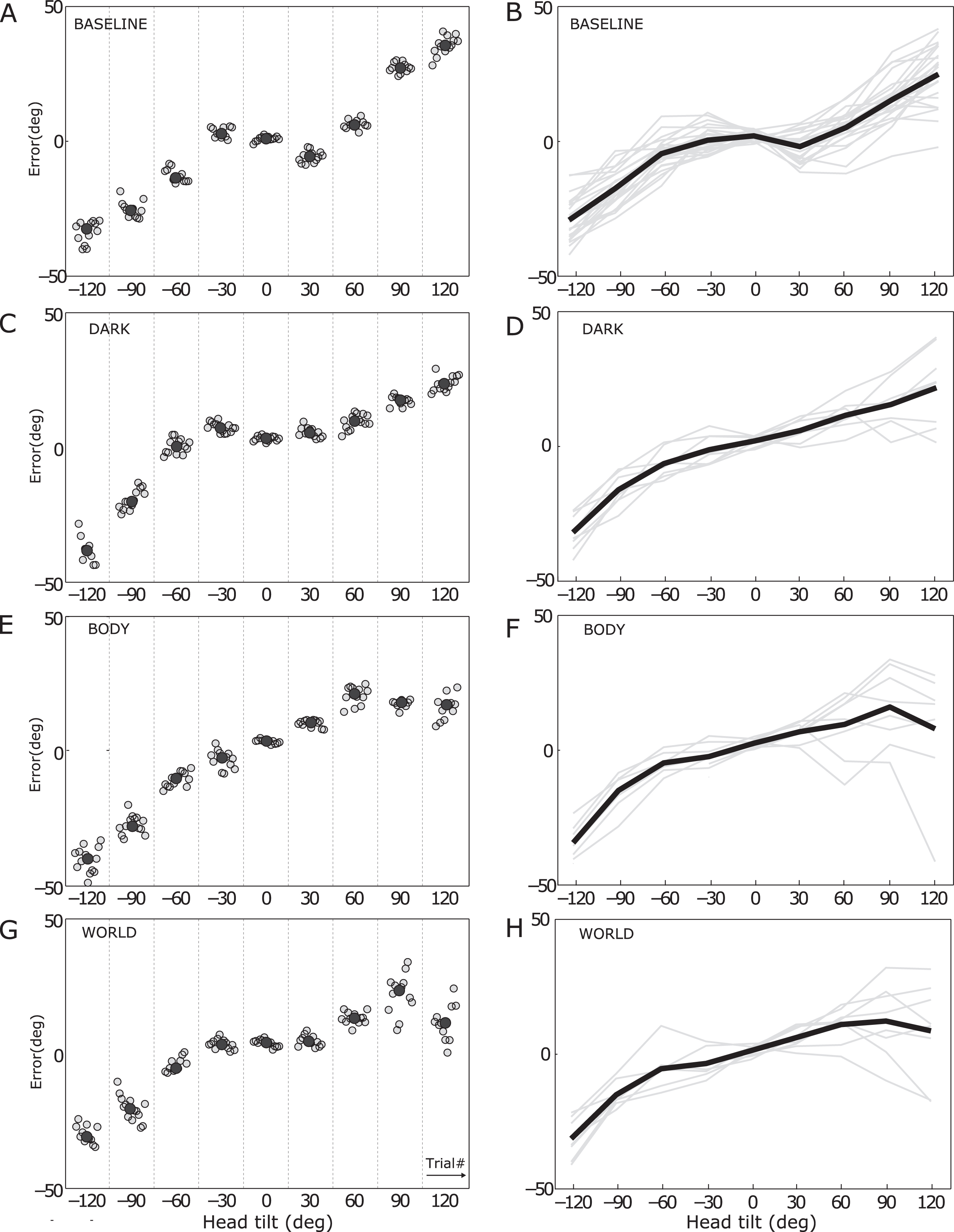
Figure 2C shows the individual adjustment errors after dark adaptation, for the same subject as in Fig. 2A-B. At tilt angles 30° and 60°, errors were shifted by about 10° toward the adaptation angle (90°), compared to the baseline data. This confined distortion of the SVV after the adaptation is seen in all individual subjects (Fig. 2D) and is consistent with previous observations [29]. Figures 2E and 2G show the individual SVV errors after adaptation with body- and world-aligned panoramic cues, respectively. The errors of the example subject look very similar for the world and body condition; the same is seen in the average response curves across all subjects (see Fig. 2F and H).
For further visual evaluation, Fig. 3 superimposes the mean error patterns across subjects in the four conditions. A complex pattern of differences emerges: at 30° and 60° tilt a clear difference is visible between the three adaptation conditions and the baseline condition, but not between the adaptation conditions. At 120° tilt, the two adaptation conditions with panoramic cues (World and Body conditions) show a clear reduction of the SVV error compared to the baseline and dark condition. The effects were asymmetric, i.e., only observed ipsilateral to the adapted tilt angle. At 90° tilt angle, all conditions yielded similar results.
Fig. 3
Effect of adaptation. SVV error (mean and SE across participants) the three adaptation conditions (gray) and the baseline (black). At 30° tilt, the error in the three adaptation conditions is reverted with respect to the baseline condition. The presence of visual panoramic cues during the tilt adaptation (World and Body conditions) also yields a reduction of the SVV error at 120° to the side of adaptation.
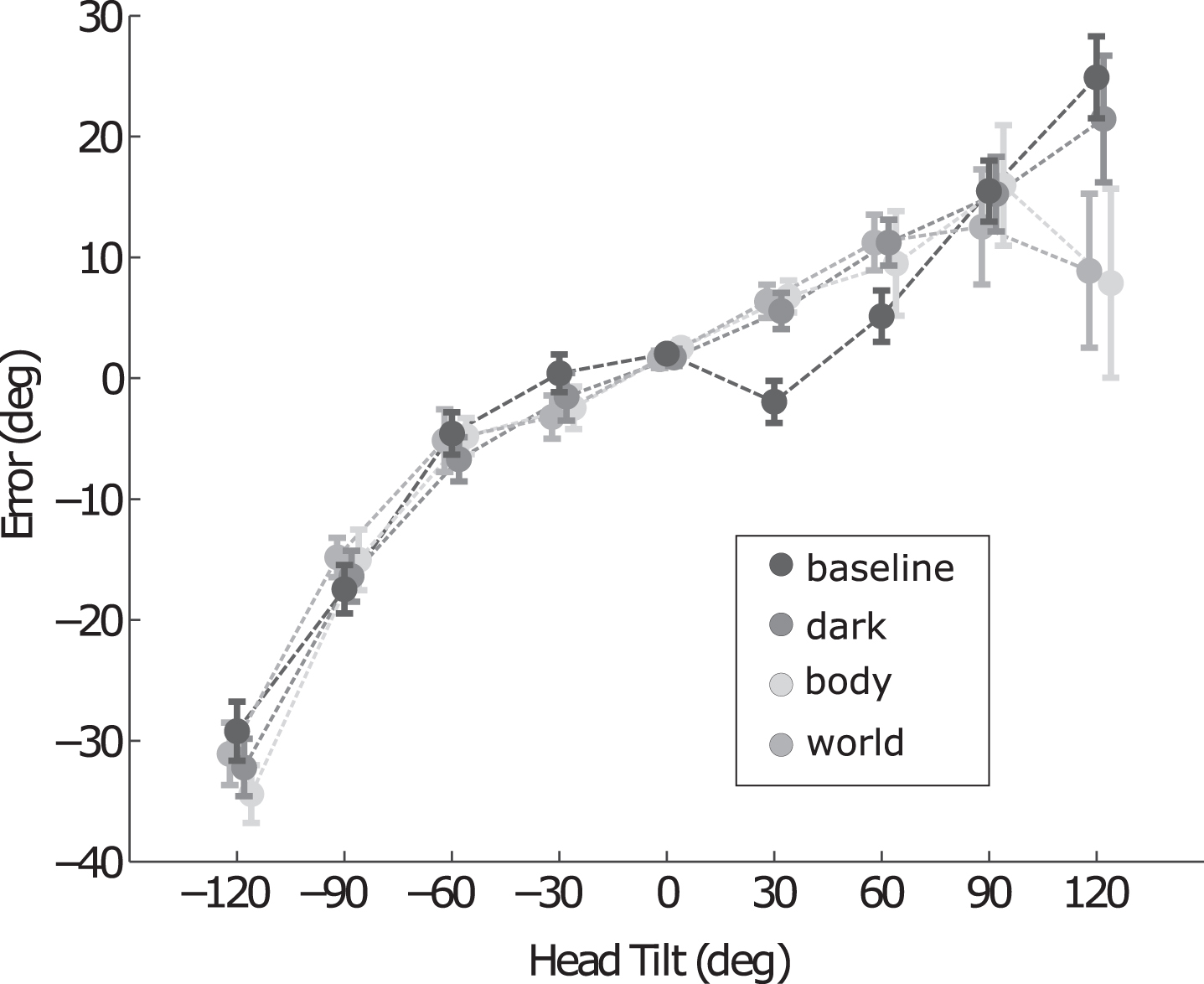
To statistically evaluate these observations, we split the analyses for clockwise and counterclockwise tilt angles. While an ANOVA showed no interaction effect between tilt angle and the four experimental conditions when the subject is tilted counterclockwise (negative tilt angles; F(7,12) =1.71, p = 0.08), this interaction is significant for roll tilts to the side of adaptation (F(7,12) =8.95, p < 0.01). To further unravel this interaction, we ran separate ANOVAs with the factor condition for the individual tilt angles. We found significant differences between conditions only at 30° (F(7,3) =12.45, p < 0.01) and 120° (F(7,3) =8.98, p < 0.01). Subsequent paired t-tests showed that in the dark condition the sign of the SVV is reverted with respect to the baseline at 30° (Baseline: –1.95°±1.73°, Dark: 5.57°±1.49°, mean±SE, p < 0.01). This effect was also found in both the body (Baseline: –1.95°±1.73°, Body: 6.76°±1.32°, p < 0.01) and world conditions (Baseline: –1.95°±1.73°, World: 6.37°±1.37°, p < 0.01). Furthermore, compared to the dark condition, the body and world conditions revealed a systematic reduction of the SVV at 120° clockwise tilt (Body: 7.86°±7.82°, Dark: 21.45°±5.24°, p < 0.04; World: 8.88°±6.37°, Dark: 21.45°±5.24°, p < 0.01). Finally, we explicitly tested for differences between the body and world adaptation conditions using a two-factor ANOVA (condition x angle). This test revealed no main effect of condition (F(1,21) = 0.08, p = 0.79) or an interaction with tilt angle (F(1,84) = 0.76, p = 0.56). This suggest that the panoramic cues modulate the SVV due to their effect on head orientation.
4Discussion
We investigated the influence of panoramic visual cues, presented during 4-minutes of static roll-tilt at 90° (clockwise tilt), on the subsequent visual assessment of the gravitational vertical (world and body conditions). For comparison, we also tested the SVV after adaptation, but without accompanying panoramic cues (dark condition), and the SVV without any prior adaptation (baseline condition).
The baseline condition demonstrated the classical pattern of results. The SVV is veridical when participants are upright, but is biased when tilted. Based on this bias, it seems that roll tilt is overestimated for small roll angles (< 60°), known as the E-effect [23], and underestimated for larger roll tilts (> 60°), known as the A-effect [2]. We also replicated the findings of Tarnutzer et al. (2014) showing that prolonged static roll-tilt in darkness distorts the subsequent SVV at small tilt angles ipsilateral to the adaptation position [29]. More specifically, SVV errors measured at 30° and 60° tilt angles shifted toward the values found at 90° tilt in the baseline condition.
Our results also indicate that visual panoramic cues play a role in the adaptation process. After subjects viewed panoramic cues during adaptation, there was additional distortion compared to the other conditions at 120° tilt, the direction of which was independent the polarization axis of the panoramic image. The local distortions, similar to those observed by Tarnutzer et al. (2014) at 30° and 60° [29], were unaffected by the presence of the visual cues during adaptation. In the following discussion of the present results, we rely on the assumption that adaptation has occurred during the roll tilt, i.e. the SVV biases foster an indirect effect of the adaptation period. A further (physiological) assessment of the adaptation effect during sustained viewing of visual, panoramic cues would be an important quest for future studies.
The presence of an observable effect only at 120° tilt may imply that the vestibular and visual-induced adaptation effects operate by separate mechanisms. In fact, it seems as if the panoramic cue does not impact the effect of adaptation on the E-effect range, but does so where the A-effect is typically observed. In the literature, the E-effect has been associated with uncompensated ocular counterroll [5, 7, 35]. If the vestibular, but not visual, adaptation affects the E-effect, it may suggest that the OCR in our study is mostly vestibular-driven (see also Fig. 1 in Pomante et al. 2019 [25]). In support, Otero-Millan and Kheradmand (2016) showed no significant correlation between the drifts in upright perception and ocular torsion during a static head tilt [24]. Following the same reasoning, one could infer that the A-effect is based on a supramodal head orientation signal, derived as an aggregate of multiple sensory cues. Of note, similar to Tarnutzer et al. (2014), we did not find systematic adaptation effects on the SVV at the adaptation position itself [29]. These authors speculated that the previous adaptation position becomes the new reference position in a strategy to counteract the growth of roll undercompensation near the most recent sustained stable head-roll position.
The second characteristic of our results is that the same distortion was found irrespective of whether the panoramic cue was aligned to the longitudinal axis of the body or to the direction of gravity. This could be taken to suggest that panoramic cues cause a local (or confined) adaptation of head orientation, which due to the 90° periodicity of their polarization effects, bears out as an additional bias on the percept of vertical. Hence, contrary to the first hypothesis presented in the Introduction, sustained exposure to conflicting panoramic and vestibular verticality cues does not result in adaptation of the perceptual interpretation of the two cues toward each other. If panoramic cues during prolonged tilt would be used to re-calibrate our sense of vertical according to the orientation of the visual scene, we should have found opposite effects in the two panoramic conditions. A gravity-aligned panoramic cue should have pulled the SVV toward the polarization direction of the visual cue, aligned to the true vertical. This should have resulted in a generalized reduction of the SVV error across all tilt angles, compared to both the dark condition and to the baseline. Our data show such a reduction only at 120° tilt. Likewise, the hypothesis of a global re-calibration would predict the body-aligned panoramic cue (body condition) during the adaptation to pull the SVV toward the body axis. This is not seen in our data, and even opposite to the observed SVV changes at 120° tilt. Thus, taken together, our hypothesis that the panoramic cues re-calibrate our sense of vertical toward their polarization direction is inconsistent with our experimental findings.
The hypothesis predicting that both panoramic visual cues induce the same post-adaptation effects on the SVV is supported by our data. The reduction of the error at 120° tilt, and the shift of the bias at 30° and 60° tilt, create a symmetric effect around the 90° adaptation position that might reveal an adaptation mechanism which pulls the head and body orientation percept toward the adapted angle. This hypothesis, however, would imply also a compression on both sides in the dark condition, which is not observed in our and Tarnutzer’s (2014) data. It could be speculated that the presence of panoramic cues during adaptation affected the sense of spatial awareness. For example, Kaptein and Van Gisbergen have suggested that at large tilt angles (135° – 225°), the lack of spatial awareness causes subjects to either align the SVV to the direction of their feet or to the perceived upright [16, 17]. This is typically expressed as a bimodal response distribution. The large variability across subjects at 120° tilt in panoramic conditions may be indicative of a similar phenomenon (see Figs. 2F and 2H), as if the bi-stable zone was shifted. The panoramic cues may shift the bifurcation tilt angle from a single to a bi-stable reference toward the adapted position. Yet, this interpretation should be interpreted with care since responses across subjects appear also more variable at –120° tilt, as if the increased variability is due to being inverted. Unfortunately, we cannot further verify these explanations without additional measurements.
Our preferred explanation for that post-adaptation, SVV is similar for the two panoramic visual cues is that these cues mediate their effects indirectly, through a global visual mechanism that interprets a panoramic cue as a head orientation cue [20]. It has been argued that panoramic cues have the same informative content for head orientation if they are rotated by 90° [1, 22, 32], although this is based on their presence during the SVV task, not during a prior stage of tilt adaptation. However, if panoramic images are cues to head orientation, and if prolonged exposure to these cues causes adaptation of head orientation, then it is most plausible that the adaptation effects bear out in the same direction, as is implied by our second hypothesis, and supplement the vestibular adaptation effects on head orientation
Acknowledgments
The research leading to these results has received funding from the European Union Seventh Framework Programme (FP7/2007-2013) under grant agreement no. 604063 (AP, WPM). The work has also been supported by the Netherlands Organization for Scientific Research, no. NWO-VICI 453-11-001 (WPM).
References
[1] | Alberts B.B.G.T. , de A.J. , Brouwer L.P.J. , Selen and and Medendorp W.P. , A Bayesian Account of Visual-Vestibular Interactions in the Rod-and-Frame Task, ENeuro 3: ((2016) ). |
[2] | Aubert H. , Eine scheinbare bedeutende Drehung von Objecten bei Neigung des Kopfes nach rechts oder links, Archiv Fur Pathologische Anatomie Und Physiologie Und Fur Klinische Medicin 20: ((1861) ), 381–393. |
[3] | Brown E.L. , Hecht H. and Young L.R. , Sensorimotor aspects of high-speed artificial gravity: I. Sensory conflict in vestibular adaptation, J Vestib Res 12: ((2002) ), 271–282. |
[4] | Carriot J. , Jamali M. and Cullen K.E. , Rapid adaptation of multisensory integration in vestibular pathways, Front Syst Neurosci 9: ((2015) ), 59. |
[5] | Clemens I.A.H. , de M. , Vrijer L.P.J. , Selen J.A.M. , van Gisbergen and Medendorp W.P. , Multisensory processing in spatial orientation: An inverse probabilistic approach, Journal of Neuroscience 31: ((2011) ), 5365–5377. |
[6] | Clément G. and Ngo-Anh J.T. , Space physiology II: adaptation of the central nervous system to space flight–past, current, and future studies, Eur J Appl Physiol 113: ((2013) ), 1655–1672. |
[7] | De M. , Vrijer W.P. , Medendorp and Van Gisbergen, J.A.M. , Accuracy-precision trade-off in visual orientation constancy, Journal of Vision 9: ((2009) ), 1–15. |
[8] | Dyde R.T. , Jenkin M.R. and Harris L.R. , The subjective visual vertical and the perceptual upright, Exp Brain Res 173: ((2006) ), 612–622. |
[9] | Eron J.N. , Cohen B. , Raphan T. and Yakushin S.B. , Adaptation of orientation vectors of otolith-related central vestibular neurons to gravity, J Neurophysiol 100: ((2008) ), 1686–1690. |
[10] | Eron J.N. , Cohen B. , Raphan T. and Yakushin S.B. , Adaptation of orientation of central otolith-only neurons, Ann N Y Acad Sci 1164: ((2009) ), 367–371. |
[11] | Fernandez C. and Goldberg J. , Physiology of peripheral neurons innervating otolith organs of the squirrel monkey. I. Response to static tilts and to long-duration centrifugal force, J Neurophysiol 39: ((1976) ), 970–984. |
[12] | Garrick-Bethell I. , Jarchow T. , Hecht H. and Young L.R. , Vestibular adaptation to centrifugation does not transfer across planes of head rotation, J Vestib Res 18: ((2008) ), 25–37. |
[13] | Gauthier G.M. and Robinson D.A. , Adaptation of the human vestibuloocular reflex to magnifying lenses, Brain Res 92: ((1975) ), 331–335. |
[14] | Hallgren E. , Kornilova L. , Fransen E. , Glukhikh D. , Moore S.T. , Clément G. , Van Ombergen A. , MacDougall H. , Naumov I. and Wuyts F.L. , , Decreased otolith-mediated vestibular response in 25 astronauts induced by long-duration spaceflight, J Neurophysiol 115: ((2016) ), 3045–3051. |
[15] | Higashiyama A. and Koga K. , Perceived self-tilt in dynamic visual stimuli: evidence for suppression by vestibulo-tactile inputs, Perception 38: ((2009) ), 281–291. |
[16] | Kaptein R.G. and Van J.A.M. , Gisbergen, Interpretation of a discontinuity in the sense of verticality at large body tilt, J Neurophysiol 91: ((2004) ), 2205–2214. |
[17] | Kaptein R.G. and Van J.A.M. , Gisbergen, Nature of the transition between two modes of external space perception in tilted subjects, J Neurophysiol 93: ((2005) ), 3356–3369. |
[18] | Lackner J.R. and DiZio P. , Space adaptation syndrome: multiple etiological factors and individual differences, J Wash Acad Sci 81: ((1991) ), 89–100. |
[19] | Lechner-Steinleitner S. , Interaction of labyrinthine and somatoreceptor inputs as determinants of the subjective vertical, Psychol Res 40: ((1978) ), 65–76. |
[20] | Li W. and Matin L. , Visually perceived vertical (VPV): induced changes in orientation by 1-line and 2-line roll-tilted and pitched visual fields, Vision Research 45: ((2005) ), 2037–2057. |
[21] | Mittelstaedt H. , A new solution to the problem of the subjective vertical, Naturwissenschaften 70: ((1983) ), 272–281. |
[22] | Mittelstaedt H. , The subjective vertical as a function of visual and extraretinal cues, Acta Psychol (Amst) 63: ((1986) ), 63–85. |
[23] | Müller G.E. , Über das Aubertsche Phänomen, Z Zinnesphysiol 49: ((1916) ), 109–246. |
[24] | Otero-Millan J. and Kheradmand A. , Upright Perception and Ocular Torsion Change Independently during Head Tilt, Front Hum Neurosci 10: ((2016) ), 573. |
[25] | Pomante A. , Selen L.P.J. and Medendorp W.P. , Visual orientation uncertainty in the rod-and-frame illusion, J Vis 19: ((2019) ), 19. |
[26] | Schöne H. and De H.U. , Haes, Perception of gravity-vertical as a function of head and trunk position, Z Vergl Physiol 60: ((1968) ), 440–444. |
[27] | Tarnutzer A.A. , Bertolini G. , Bockisch C.J. , Straumann D. and Marti S. , Modulation of internal estimates of gravity during and after prolonged roll-tilts, PLoS One 8: ((2013) ), e78079. |
[28] | Tarnutzer A.A. , Bockisch C.J. and Straumann D. , Head roll dependent variability of subjective visual vertical and ocular counterroll, Exp Brain Res 195: ((2009) ), 621–626. |
[29] | Tarnutzer A.A. , Bockisch C.J. , Straumann D. , Marti S. and Bertolini G. , Static roll-tilt over 5 minutes locally distorts the internal estimate of direction of gravity, Journal of Neurophysiology 112: ((2014) ), 2672–2679. |
[30] | Van A.D. , Beuzekom and Van Gisbergen J.A. , Properties of the internal representation of gravity inferred from spatial-direction and body-tilt estimates, Journal of Neurophysiology 84: ((2000) ), 11–27. |
[31] | Van Ombergen A. , Van Rompaey V. , Maes L.K. , Van de Heyning P.H. and Wuyts F.L. , Mal de debarquement syndrome: a systematic review, J Neurol 263: ((2016) ), 843–854. |
[32] | Vingerhoets R.A.A. , De Vrijer M. , Van Gisbergen J.A.M. and Medendorp W.P. , , Fusion of visual and vestibular tilt cues in the perception of visual vertical, Journal of Neurophysiology 101: ((2009) ), 1321–1333. |
[33] | Wade N.J. , Visual orientation during and after lateral head, body, and trunk tilt, Percept Psychophys 3: ((1968) ), 215–219. |
[34] | Wade N.J. , Effect of prlonged tilt on visual orientation, Q J Exp Psychol 22: ((1970) ), 423–439. |
[35] | Wade S.W. and Curthoys I.S. , The effect of ocular torsional position on perception of the roll-tilt of visual stimuli, Vision Res 37: ((1997) ), 1071–1078. |
[36] | Ward B.K. , Bockisch C.J. , Caramia N. , Bertolini G. and Tarnutzer A.A. , Gravity dependence of the effect of optokinetic stimulation on the subjective visual vertical, J Neurophysiol 117: ((2017) ), 1948–1958. |
[37] | Witkin H.A. and Asch S.E. , Studies in space orientation; perception of the upright in the absence of a visual field, J Exp Psychol 38: ((1948) ), 603–614. |
[38] | Zaidel A. , Turner A.H. and Angelaki D.E. , Multisensory Calibration Is Independent of Cue Reliability, Journal of Neuroscience 31: ((2011) ), 13949–13962. |
[39] | Zupan L.H. , Merfeld D.M. and Darlot C. , Using sensory weighting to model the influence of canal, otolith and visual cues on spatial orientation and eye movements, Biological Cybernetics 86: ((2002) ), 209–230. |


