
Biofluid Markers and Tissue Biopsies Analyses for the Prodromal and Earliest Phase of Parkinson’s Disease
Abstract
The recent development of new methods to detect misfolded α-synuclein (αSyn) aggregates in biofluids and tissue biopsies in the earliest Parkinson’s disease (PD) phases is dramatically challenging the biological definition of PD. The αSyn seed amplification methods in cerebrospinal fluid (CSF) showed high sensitivity and specificity for early diagnosis of PD and Lewy bodies disorders. Several studies in isolated REM sleep behavior disorders and other at-risk populations also demonstrated a high prevalence of CSF αSyn positivity and its potential value in predicting the phenoconversion to clinically manifested diseases. Growing evidence exists for αSyn aggregates in olfactory mucosa, skin, and other tissues in subjects with PD or at-risk subjects. DOPA decarboxylase and numerous other candidates have been additionally proposed for either diagnostic or prognostic purposes in earliest PD phases. The newly described αSyn detection in blood, through its quantification in neuronally-derived exosome vesicles, represents a technical challenge that could open a new scenario for the biological diagnosis of PD. Despite this growing evidence in the field, most of method of αSyn detection and markers still need to be validated in ongoing longitudinal studies through an accurate assessment of different prodromal disease subtypes and scenarios before being definitively implemented in clinical settings.
INTRODUCTION
α-Synucleinopathies are a group of neurodegenerative conditions, including Parkinson’s disease, dementia with Lewy bodies (DLB), multiple system atrophy (MSA), and other rarer entities defined by aggregation of abnormal forms of α-Synuclein (αSyn) and other proteins in neurons and glial cells. αSyn conformational modification into misfolded oligomeric and fibrillary forms is the most consistent pathological feature of α-Synucleinopathies. The interplay of these abnormal αSyn forms with the cellular pathways and cell organelles involved in their clearance results in neuronal dysfunction and, ultimately in axonal injury and neuronal death. The recent development of new techniques able to detect and quantify αSyn in vivo in different biofluids and tissues, including cerebrospinal fluids (CSF), plasma, olfactory mucosa, and skin, is now dramatically changing the scenario of α-Synucleinopathies, prompting the discussion for a biological definition of PD.1–3 Several studies showed that subjects later diagnosed as α-Synucleinopathies show a long prodromal phase, presenting with a combination of subtle non-motor or only mild motor features.4–6 Isolated REM sleep behavior disorder (iRBD) carries the highest positive predictive value for impending α-Synucleinopathies, with an estimated risk of phenoconversion over 70% after 12 years of follow-up.7–9 Subjects with iRBD are thus of particular interest for the study of the earliest biomarkers of α-Synucleinopathies, despite the fact they represent a specific prodromal subtype of disease with a higher risk of developing cognitive deficits and autonomic dysfunction.10,11 To disentangle the complexity underpinnings prodromal α-Synucleinopathies, several longitudinal studies are now following-up subjects with subtle non-motor features such as hyposmia, autonomic dysfunction, subtle motor alterations, language impairment or individuals with positive family history for parkinsonism or carrying specific mutation associated with PD- all conditions related to a lower but consistent risk of developing α-Synucleinopathies. 10,12–14
During the last few years, exciting results from such studies suggest that αSyn alterations are detectable even during the prodromal stages and might also be a potential target of disease-modifying intervention strategies.15 This work will review the candidate biological markers for diagnosing α-Synucleinopathies, with a special focus on fewer studies in prodromal or earliest disease phases. The review will also include potential markers discussed within the research community for the differentiation of different α-Synucleinopathies and as markers of disease progression. Particular attention is paid to the recent development of seed amplification assays targeting the abnormal forms of αSyn in vivo and the clinical application of this assay which has dramatically changed the scenario of the ideal biological marker.7,8,16 Clinical research also highlighted the value of numerous other candidates other than αSyn, including DOPA decarboxylase, neuronal, glial, and inflammatory disease associated biomarkers already in the early PD phases. The role of these biomarkers for the differential diagnosis of different α-Synucleinopathies and as possible proxies of progression and, thus, ideal targets for the disease-modifying strategies will briefly be discussed (see Fig. 1 andTable 1).
Fig. 1
Potential biofluid and tissue markers of α-Synucleinopathies. Markers have been differentiated based on high sensitivity (H) when the mean diagnostic accuracy of the markers is > 80%, I intermediate accuracy when < 80%. (P) indicated prognostic markers. DD, differential diagnosis; NfL, neurofibrillary light chain; SAA seed amplification assay.
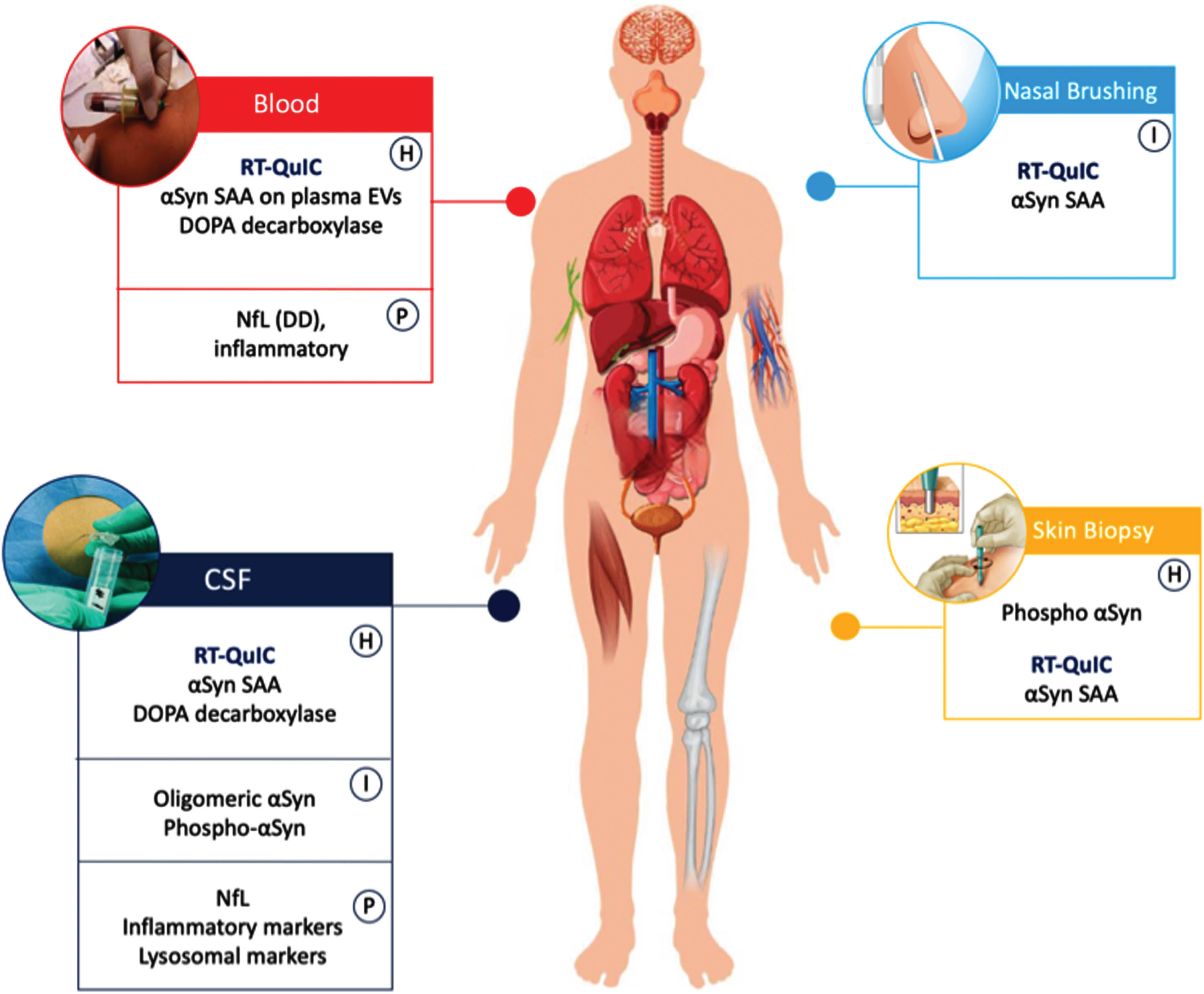
Table 1
Most important markers used in earliest phase of PD and at-risk populations
| Technique | Biological | Sensitivity | Specificity | Application in prodromal stages | ||
| Sample | Clinically Overt | Prodromal | Clinically Overt | Prodromal | ||
| oligomeric αSyn (ELISA) | CSF | moderate | low | moderate | moderate | CS &L - |
| Blood | low | NE | moderate | NE | NE | |
| RT-QuIC αSyn | CSF | high | moderate | high | high | CS &L + |
| Skin | high | intermediate | high | high | CS &L + | |
| Olfactory Mucosa | low | low | high | high | CS &L +/- | |
| Plasma EVs | high | intermediate | high | high | CS &L + | |
| Total αSyn | CSF | intermediate | NE | Low | NE | CS &L - |
| NfL (exclusion criteria, suggestive of non-PD α-Synucleinopathies) | CSF | high | NE | high | NE | CS &L - |
| Blood | intermediate | intermediate | high | high | CS &L + | |
| DOPA decarboxylase (marker of dopaminergic loss) | CSF | high | intermediate | high | high | CS &L + |
| Blood | high | intermediate | high | high | CS + | |
Abbreviations: NfL, neurofibrillary light chain; SAA, seed amplification assay, αSyn, α-Synuclein; NE: not enough evidences for the use of the specific biomarker (based on negative or inclusive findings or meta-analyses); CS: cross sectional studies; L: longitudinal studies.
– = No efficacy (also scored if negative in a meta-analysis).
+ = Efficacy in at least one study.
*High sensitivity and specificity: >80%; moderate sensitivity and specificity: >70% and ≤80%; low sensitivity and specificity: ≤70%.
αSYN DETECTION IN CSF USING STANDARD ASSAYS
Total, phosphorylated, and oligomeric αSyn levels have been explored mainly in CSF of α-Synucleinopathies, with different methods and still debated results.17
Several studies tested the correlation between total CSF αSyn concentrations in CSF and PD disease stage, but results are conflicting, with some studies reporting lower levels of αSyn in CSF of PD patients compared to matched controls while others report no differences or even higher levels of αSyn in PD cohorts.17 A first meta-analysis conducted by Zhou and coauthors in 2015 found lower mean total αSyn concentration in CSF of PD patients compared to controls and significantly higher in PD compared to MSA, with no differences between PD and DLB.18 Another updated meta-analysis found no differences in total CSF αSyn levels between PD and other parkinsonisms.19 In contrast, total αSyn was significantly reduced in PD compared to controls, yielding an estimated sensitivity and specificity of 72% and 65% respectively. The overall heterogeneity remained high even after considering blood contamination, disease staging, and duration.17 Moreover, reports show extensive signal overlap among tested individuals, diminishing its potential use in clinical practice.20 ELISA assay settings for total αSyn were quite similar among the studies. However, significant analytical differences existed in the type of antibodies, calibrators, and quantitation methods, explaining an essential source of resultsheterogeneity.21
Phosphorylated forms of αSyn have also been assessed in CSF, resulting in still conflicting results. In most studies, αSyn at serine-129 (Ser-129p-αSyn) levels have been elevated in the CSF of PD patients. However, similar increases are seen in atypical parkinsonism, limiting their diagnostic accuracy in clinical practice.17,22 Different sites of αSyn phosphorylation has been claimed to be specific for α-Synucleinopathies, but their role has been no further confirmed in more extensive validation studies.23,24
Other studies tested oligomeric αSyn in CSF as a potential marker, disclosing higher levels of PD in comparison to controls. The oligomeric/total αSyn ratio in CSF is one of the most promising biomarkers for distinguishing PD patients from controls, albeit a considerable overlap with controls and other neurodegenerative conditions was observed.25 In general, standard measures of αSyn in fluids have given inconsistent results especially when trying to differentiate between PD and atypical parkinsonisms such as MSA.
Very few studies on CSF markers have been conducted in at-risk individuals.
Aasly and coauthors found that levels of CSF αSyn oligomers were significantly elevated in healthy asymptomatic LRRK2 mutation carriers compared to controls.26 These findings were subsequently confirmed by Majbour and coauthors in an independent cohort of LRRK2 carriers, suggesting that the ratio between total and oligomeric αSyn is a risk biomarker candidate for the detection of pathological changes at the prodromal stage.27 In iRBD, Wang and coauthors reported a reduction of total αSyn levels in CSF in these patients; of note, the subjects with lower levels exhibited an increased risk of phenoconversion to α-Synucleinopathies in longitudinal analyses.28
OTHER PROMISING CSF MARKERS
The recent development of high-sensitivity techniques for CSF and plasma enables the quantification of other essential markers in CSF with potential value for differential diagnosis, subtyping, and prognosis in α-Synucleinopathies. One of the most established markers tested in neurodegeneration is the neurofilament light chain (NfL), which correlates to axonal damage and neuronal dysfunction and can be measured also in blood.29 In PD, NfL levels in CSF were investigated in several cross-sectional and longitudinal cohorts of de novo PD patients, demonstrating the highest levels in other neurodegenerative diseases and atypical parkinsonism, including MSA and DLB and atypical tauopathies such as progressive supranuclear palsy and corticobasal syndrome. Thus, CSF NfL could be used for differentiating PD from atypical parkinsonism or identify PD subtypes of disease with a more severe motor and cognitive progression subjects.30–36
In a recent paper under review, Hoglinger and colleagues suggested that elevated NfL in αSyn-positive patients is more suggestive of MSA and should be used as an exclusion criterion for Parkinson’s disease.37 In MSA patients, NfL levels are higher than in PD patients and HC both in CSF and in plasma samples 38,39 and correlated with disease severity and progression rates.40
MITOCHONDRIAL AND LYSOSOMAL MARKERS
Mitochondrial dysfunction contributes to the pathogenesis of PD.41 The most studied mitochondrial biomarker is CSF DJ-1, which shows decreased levels in PD compared to controls and it correlated with disease severity. However, similar values in other parkinsonian syndromes limit its diagnostic use.42,43
A dysregulated autophagy–lysosomal system can cause a reduced degradation of intracellular protein, thus enhancing αSyn accumulation in PD.44 This hypothesis has been confirmed by identifying mutations in the GBA gene, encoding the lysosomal enzyme glucocerebrosidase (GCase), as the most common genetic risk factor for PD 45. CSF GCase activity depends on the specific GBA1 mutations, but low levels are also reported in idiopathic PD patients compared with controls.46 GCase levels are, however, of low value for diagnosing PD, and moderate diagnostic accuracy of lysosomal enzymes can only be yielded in combination with other markers; GCase activity, the o-/t- αSyn ratio, and age showed the best performance in discriminating PD from controls independent of GBA mutation status.47 Moreover, CSF GCase levels correlate with cognitive impairment 48 also in idiopathic PD, underlying the potential role of lysosomal biomarkers in stratifying subjects with different disease subtypes. In contrast, no consistent data in prodromal phases has still been published.
DOPA-DECARBOXYLASE
DOPA-decarboxylase (DDC), an essential enzyme in neurotransmitter metabolism, has been recently suggested as a promising marker by two independent publications using large-scale multiplex proximity extension assay (PEA) on the Olink platform.49,50 Findings showed an elevation of DDC in both CSF and plasma of people with PD or atypical parkinsonism and has been shown to be a biomarker of dopaminergic cell loss.51 DDC levels in CSF can accurately identify patients with Lewy body disease (LBD) (AUC = 0.89 in Pereira et al.50, 0.80 in Paslawski et al.49) and are associated with worse cognitive performance. Furthermore, DDC can detect preclinical LBD stages in asymptomatic individuals with a positive seed amplification α-synuclein assay (AUC = 0.81 in Pereira et al.50) and that this biomarker could predict progression to clinical LBD over a 3-year period.50
NEUROINFLAMMATORY CSF MARKERS
Mild neuroinflammation plays a major role in the pathology of PD since αSyn triggers the activation of the immune system52 and inflammatory mediators have been investigated in CSF as candidate biomarkers for PD.
Neutrophil-to-lymphocyte ratios (NLRs), related to the overall inflammatory status, and glial fibrillary acidic protein (GFAP), a protein released by astrocytes, have been potentially suggested by recent publications as able to distinguish PD from healthy controls, despite a large overlaps of individual values and lack of significance of other studies.15,49,50,53 The already cited study of Paslawski and coauthors identified Midkine (MK) using the PEA approach as specifically increased in α-Synucleinopathies (AUC 0.78). In the same study, the findings were validated using ELISA and correlated with the anti-inflammatory cytokines IL- 17D and IL-27cytokines expressed by astrocytes and has been suggested to play a role in local immune responses.49 Other markers can reliably discriminate tauopathies from α-Synucleinopathies such as YKL-40 (chitinase-3-like protein 1) and MCP-1 (monocyte chemoattractant protein-1);54,55 however, current evidence does not suggest that these markers would help in distinguishing clinically manifested PD from its earliest phases and from atypical conditions, also due to limited numbers of studies available.15 Myelin basic protein (MBP) could represent a promising biomarker for MSA, considering that demyelination is a key features of MSA pathogenesis. Indeed, some studies have showed higher CSF levels of MBP in MSA patients compared to PD patients and HC.56,57
In at-risk populations, the limited number of subjects who underwent CSF still limited its study in large longitudinal cohorts, and the recent development of blood-based assays represents a significant step forward for a wider use of biological markers in the earliest phases of α-Synucleinopathies.
SEED AMPLIFICATION ASSAYS AND αSYN DETECTION IN CSF
Recently, it has been shown that misfolded αSyn aggregates adopt alternative conformations and become self-propagating as prion proteins causing the rare Creutzfeldt-Jakob disease in humans.58 Protein misfolding cyclic amplification (PMCA) and Real-Time Quaking-induced conversion (RT-QuIC) assays exploited this self-replicating nature of prions, approaching 100% sensitivity and specificity by applying this in human prion disorders.59,60 To avoid confusion between RT-QuIC assay for prions and for other misfolded proteins, the consensus name ‘seed amplification assay (SAA)’ has emerged to refer to non-prion assays that exploit self-replication of misfolded proteins by means of fragmentation and elongation cycles, such as αSyn, tau, and TDP-43.61–63
Several studies confirmed excellent sensibility and sensitivity of αSyn SAA in differentiating healthy controls from clinically established PD patients in CSF samples.16,64 Many studies have shown that αSyn SAA has a sensitivity and specificity above 90% for PD and for other α-Synucleinopathies.64,65 The technique showed very high specificity when αSyn SAA was analysed in CSF from neuropathologically confirmed cases.65 A recent meta-analysis on 22 SAA studies in CSF showed an overall sensitivity of 0.88 (95% CI, 0.82–0.93) and specificity of 0.95 (95% CI, 0.92–0.97) for distinguishing α-Synucleinopathies versus non α-Synucleinopathies including tauopathies.64 In the recent study published on the PPMI cohort by Siderowf and colleagues, the sensitivity for established PD resulted in 87.7% with a specificity for healthy controls of 96.3%. Of note, the sensitivity increases in subjects with typical olfactory deficits up to 98.6% and decreases to 78.3% in those without hyposmia and 67.5% in LRRK2 mutation carriers. This fits the assumption that αSyn SAA in CSF might be associated with specific disease subtypes and mutations, including GBA,66 with even possible prognostic implications.
In DLB, an early meta-analysis showed a sensitivity of 0.95 and a sensibility of 0.96 for CSF samples alone 67 compared to controls. In patients with dementia, however, the common detection of subjects with biologically established Alzheimer’s disease but positive αSyn SAA should be considered as a possible diagnostic challenge, especially in older subjects.68
In MSA, the application of seed aggregation assays provided variable and inconsistent results: some studies showed a good performance in detecting αSyn in MSA patients, while some found very low rates of RT-QuIC positive MSA patients. This inconsistency could be partially due to inter-laboratory experimental parameters variability, but also to claimed different conformation of misfolded αSyn in MSA compared to either PD and DLB.69 The kinetics parameters of RT-QuIC could also differentiate MSA from PD and DLB in some of these studies.70 This might explain the prominent differences in sensitivity (0.30 for αSyn SAA specifically) detected in MSA patients compared to PD. Interestingly, some studies have demonstrated exemplary performance in combining the extent of αSyn seeding activity with NfL levels to distinguish MSA from PD.71 In one study, published by Poggiolini et al., RT-QuIC parameters was also associated with rate of progression in MSA patients.8
SAA IN AT-RISK AND PRODROMAL POPULATIONS
Only a few studies evaluated αSyn SAA seeding activity in CSF of subjects with prodromal PD, mainly including iRBD and genetic carriers.64 Considering the whole studies included in the meta-analyses (which unfortunately excluded a few important recently published works), αSyn-SAAs yielded a pooled sensitivity and specificity to differentiate patients with prodromal signs of α-Synucle-inopathies from non-α-Synucleinopathies of 0.74 (95% CI, 0.36–0.93) and 0.93 (95% CI, 0.89–0.96), respectively in these phases of the disease. The diagnostic performance of αSyn-SAAs for patients with RBD showed sensitivity rates of 0.64 (95% CI, 0.50–0.77)8 and 0.80 (95% CI, 0.58–0.92)72 and 1.00 (95% CI, 0.82–1.00).65 Pooled sensitivity and specificity rates could not be computed for this analysis due to the small number of included studies. Notably, the recently published cross-sectional study evaluating the PPMI at-risk groups indicated a sensitivity of SAA assays in 86% of subjects with iRBD/hyposmia and 8% of mutation carriers.16
Unfortunately, only a few studies included longitudinal assessment and revision of conversion to clinical α-Synucleinopathies. The longitudinal study of Iranzo and co-authors found a diagnostic accuracy of 90% in predicting the phenoconversion of iRBD, considering a follow-up up to ten years.72 A recent study found 92% positivity in subjects with pure autonomic failure. This study showed a crucial prognostic value of the magnitude and reaction kinetic of SAA in predicting the time to conversion to established α-Synucleinopathies.73 No extensive longitudinal studies are available on subjects with hyposmia or mutation carriers.
SAA IN OLFACTORY NEUROEPITHELIUM
The olfactory neuroepithelium (OE) is a neural tissue mainly located in the upper nasal cavity. It is directly exposed to the external environment and vulnerable to physical and chemical injuries. OE comprises various cell types, including olfactory sensory neurons, supporting glial-like cells, microvillar cells, and basal stem cells. Nasal brushing (NB) was initially set up to collect OE neural cells for prion disorders diagnosis. It is a non-invasive procedure and easy to be performed without medical contraindications or therapy restrictions. In patients with sporadic Creutzfeldt-Jakob disease, the pathologic prion protein was detected in olfactory mucosa by Real-Time Quaking-Induced Conversion (RT-QuIC), yielding a diagnostic accuracy of nearly 100%.74,75
In neurodegenerative disorders such as AD or PD, different studies have shown the presence of neurofibrillary tangles, amyloid-β deposits, or Lewy neurites in the olfactory sensory neurons (ONs).76,77 Subsequently, we demonstrated that OE expressed all proteins associated to different neurodegenerative disorders, such as αSyn, TDP-43, tau, prion protein, APP indicating that the pathological aggregated forms, linked to specific proteinopathy, might be potentially detected in this neural tissue.62,78
Thus, in subsequent studies, NB has been applied to patients with PD, DLB and to patients with iRBD for detecting αSyn aggregates by αSyn SAA. Among these αSyn-related disorders, a variable αSyn SAA sensitivity ranged from ∼80% in DLB to ∼45% in PD and iRBD was observed.79 Since NB was usually performed at the level of middle turbinate (mt), NB was performed from an olfactory area with a higher concentration of olfactory neurons such as at the level of upper turbinate (agger nasi). αSyn SAA sensitivity increased from 45% to 80% if NB was performed at the level of upper turbinate instead of middle turbinate, and this difference correlated to the higher concentration of olfactory neural cells at the level of agger nasi.74 In addition, in OE from PD patients but not in controls, an increased expression of αSyn and phospho-αSyn deposits were observed. PD and MSA patients showed higher seeding activity than HC and other neurodegenerative diseases; moreover, also in OE αSyn strains morphology differs between MSA and Lewy body pathologies.80 Intriguingly, patients with the cerebellar MSA variant did not show any seeding activity, so OE α-synuclein RT-QuIC could potentially become a biomarker to distinguish these two clinical entities.81 Although preliminary, these studies indicate that NB is a simple and harmless procedure for collecting ONs and neural cells to be tested by αSyn SAA using specific protein substrates of replication. This allows a molecular diagnosis of α-Synucleinopathy in clinically affected patients and during prodromal stages and αSyn SAA positivity might represent an essential marker of phenoconversion in addition to that detected in CSF, which appeared to be relatively stable over time. Further ongoing longitudinal studies in iRBD and other at-risk populations will definitively disentangle the application of NB coupled with SAA in α-Synucleinopathies.
TISSUE BIOPSIES
Phosphorylated αSyn (p-αSyn) deposits can be found at several central and peripheral nervous system levels in patients with α-Synucleinopathies, both in their prodromal and symptomatic phases.82
In patients with RBD, seeding of p-αSyn was first reported in colonic tissue, with a very low positivity rate (24%).83 Higher sensitivity (89%) was then obtained with submandibular gland biopsy. Still, adequate biopsy material can be obtained in less than half of the patients,84 while minor salivary glands biopsy obtained adequate tissue in all cases, although showing less sensitivity (50%).85
Lately, skin biopsy has emerged as a promising and less invasive technique. In 2017, two groups reported high sensitivity and specificity of this technique in detecting p-αSyn deposits utilizing immunofluorescence (IF) analysis. One study using biopsies of multiple unilateral sites showed a positivity of 56% in RBD patients (10 out of 18 patients) and 80% in PD patients (20/25 patients), with high specificity (0 out of 20 controls).86 A second study independently confirmed this finding using bilateral biopsies at C8 and the distal leg, showing p-αSyn positivity in 9 (75%) of 12 patients with isolated RBD and 0 of 55 controls. Merged follow-up data of these two original cohorts showed consistent findings over time (two to three years of follow-up), with phenoconversion reported only in patients with a positive skin biopsy.87 The analysis technique has shown excellent interobserver reliability in these two independent laboratories.88 Subsequent studies further confirmed these data.89,90
The distribution of pS129-αSyn varies among α-Synucleinopathies: in fact, the deposition involves more often the somatic sensory fibers in MSA, whilst in PD and DLB the involvement is prevalent in autonomic fibers.91,92
Recently, studies have focused on measuring αSyn by means of αSyn SAA. The first group compared IF and αSyn-seed amplification assay (αSyn SAA) of skin and CSF in 41 iRBD and 40 matched controls, reporting an excellent diagnostic accuracy (89%) of IF and a lower accuracy in the case of skin and CSF-based αSyn-SAA (70% and 69%, respectively).93
In another cohort, analyzed by Kuzkina et al. in 2023, αSyn aggregation using αSyn SAA was detected in 97.4% of iRBD patients (78.4% of iRBD biopsies), 87.2% of PD patients (70% of PD biopsies), and 13% of controls (7.9% of control biopsies), with a higher seeding activity in iRBD compared to PD, confirming αSyn SAA being sensitive but less specific than IF in differentiating α-Synucleinopathies since their prodromal stages.94
Iranzo compared the results of αSyn SAA in the skin and CSF in 91 patients with IRBD and 41 controls. RT-QuIC detects αSyn in the skin and CSF with high sensitivity (>75%), specificity (>97%), and similar agreement.95
Overall, searching for misfolded αSyn in the skin with IF or αSyn SAA turned out to be a sensitive and not invasive technique for providing diagnosis of α-Synucleinopathies, with IF analysis probably carrying higher specificity. One aspect that may directly impacts on the tool sensitivity is the choice of the best site of biopsy. There is some evidence of a proximal-to-distal gradient of αSyn positivity in patients with PD, but large studies systematically comparing different biopsy sites are lacking.96
Longitudinal studies are needed to better understand its full potential as a diagnostic and prognostic marker of conversion. In that regard, in a recent survey, αSyn misfolding was found less commonly in the olfactory epithelium than in the skin.97 Its distribution appeared not uniform, with a higher deposition of misfolded αSyn across all sampled tissues in the iRBD cohort compared to PD,87 allowing the author to infer that this technique also holds promises for patient stratification.
PLASMA MARKERS
During the last few years, consistent results coming from different laboratories showed that αSyn SAA detection in blood is feasible, especially when vesicles have been previously selected with neuronal-specific markers.98–100 Indeed, a growing number of experience found seeding activity from pathological αSyn derived from plasma extracellular vesicles (EVs).99 A recent report demonstrating the excellent ability of serum immunoprecipitation-based (IP) αSyn SAA to distinguish PD from healthy controls may herald a new approach to diagnosing PD via a simple blood test. However, lower detection rates in MSA, likely due to technical factors, still needs to be overcome.101
The IP/RT-QuIC method was applied to various αSyn-related disease forms including α-Synucleinopathy patients, non-α-Synucleinopathy patients, healthy individuals, familial PD patients with parkin gene mutations, and REM sleep behavior disorder patients. The results showed significant detection of αSyn seeds in α-Synucleinopathies, with high diagnostic performance for differentiating PD from controls (AUC: 0.96 (95% confidence interval (CI) 0.95–0.99). In pathologically confirmed cases, positivity rates of IP/RT-QuIC varied among different patient groups. The transmission electron microscopy analysis revealed distinct fibrillary structure of the IP/RT-QuIC-derived αSyn fibrils, as well as their propagation potential was confirmed. The structural differences of αSyn fibrils in PD and MSA also corresponded to unique morphology of αSyn intracellular inclusions. One of the reasons for the low positive rate of MSA is that the seed aggregation rate and fibril structure are different from PD. Whether the fibrils from MSA are unstable or the proper amplification conditions differ depending on the disease requires further study.
With a different technique, Kluge and colleagues showed in different independent cohorts an increased level of αSyn amplified by SAA in neuronally derived EVs.99,102 Recently, the same group re-evaluating subjects who developed PD demonstrated that all subjects exhibited a SAA positivity at time of diagnosis and that such alterations could appear 1 to 10 years before it with iRBD showing 30% of positivity.103
Several further studies assessing the neuronal marker NfL are available for PD, indicating a low to moderate increase in subjects with PD, which could be useful in the differentiation against MSA and DLB or other tauopathies such as PSP or CBS (AUC = 86%; sensitivity = 56%; specificity = 89% in differentiating PD from atypical parkinsonism, with AUC = 95%; sensitivity = 51% and specificity = 100% for CBS/PSP and AUC = 88%; sensitivity = 57% and specificity = 90% for MSA).104,105 In PD, several studies also found that the subset of subjects with higher values are associated with a higher longitudinal risk of motor progression and dementia.104,105 In DLB, NfL is associated with severity and is an important potential predictor of progression in the prodromal MCI phase.106
In prodromal PD, several studies are ongoing. GBA mutation carriers did not show any difference in evaluating ferritin and immunological markers compared to age-matched controls.107
Recently, Yan and coauthors found a high discrimination accuracy for identifying subjects with iRBD or prodromal PD markers using an independent validation design.108 However, most studies are cross-sectional and focused on specific subtypes of prodromal α-Synucleinopathies. One of the few longitudinal studies published using the SIMOA technique indicated NfL as a promising biomarker of conversion to clinically manifested α-Synucleinopathy using longitudinal testing and evaluating the individual changes over time in at-risk subjects.109 Another longitudinal study on iRBD, published by Zhang et al. in 2023, confirmed that plasma NfL is valuable in reflecting disease severity of iRBD and predicting disease progression and phenoconversion.110
Despite these inspiring results in the field, further results are needed to understand the role of blood αSyn SAA and other markers for the risk stratification of subjects at risk and in the earliest phases of α-Synucleinopathies.
CONCLUSIONS
The recent development of SAA and new sensitive assays for biomarker detection in biofluids and tissues is dramatically changing the scenario of PD diagnosis. Still, most studies are performed when the clinical diagnosis is manifested, and most of the works in prodromal/earliest phases of the diseases are cross-sectional. Especially for SAA, few laboratories have experience in this assay, and further inter-laboratories exchange and cross-validation studies are needed to validate the reliability and stability of results. In addition, the vast heterogeneity of clinical presentation of α-Synucleinopathies should be also considered for both at-risk target populations and the distribution of markers in different tissues, which might underline different pathophysiology mechanisms and subtypes of diseases. The combination of varying techniques and markers is pivotal to detecting and differentiating markers of stage and progression, which are particularly interesting as target engagement for disease-modifying treatments and strategies.
The use of biological markers for diagnosing PD and α-Synucleinopathies even before the development of symptoms is a significant challenge for the research and clinical community. Still, it will definitively change the scenario of PD clinical approach in the subsequent decades.
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
The authors have no funding to report.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
Nobutaka Hattori is part of the Editorial Board Member of this journal but was not involved in the peer-review process of this article nor had access to any information regarding its peer-review
REFERENCES
1. | Cardoso F , Goetz CG , Mestre TA , et al. A statement of the MDS on biological definition, staging, and classification of Parkinson’s disease. Mov Disord. (2024) ; 39: : 259–266. |
2. | Chahine LM , Merchant K , Siderowf A , et al. Proposal for a biologic staging system of Parkinson’s disease. J Parkinsons Dis. (2023) ; 13: : 297–309. |
3. | Höglinger GU , Adler CH , Berg D , et al. Towards a biological definition of Parkinson’s disease. Preprints. (2023) ; https://doi.org/10.20944/preprints202304.0108.v1 [Preprint]. Posted April 7, 2023. |
4. | Heinzel S , Berg D , Gasser T , et al. Update of the MDS research criteria for prodromal Parkinson’s disease. Mov Disord. (2019) ; 34: : 1464–1470. |
5. | Pilotto A , Heinzel S , Suenkel U , et al. Application of the movement disorder society prodromal Parkinson’s disease research criteria in 2 independent prospective cohorts. Mov Disord. (2017) ; 32: : 1025–1034. |
6. | Yilmaz R , Suenkel U , Postuma RB , et al. Comparing the two prodromal Parkinson’s disease research criteria-lessons for future studies. Mov Disord. (2021) ; 36: : 1731–1732. |
7. | Miglis MG , Adler CH , Antelmi E , et al. Biomarkers of conversion to α-synucleinopathy in isolated rapid-eye-movement sleep behaviour disorder. Lancet Neurol. (2021) ; 20: : 671–684. |
8. | Poggiolini I , Gupta V , Lawton M , et al. Diagnostic value of cerebrospinal fluid alpha-synuclein seed quantification in synucleinopathies. Brain. (2022) ; 145: : 584–595. |
9. | Postuma RB , Iranzo A , Hu M , et al. Risk and predictors of dementia and parkinsonism in idiopathic REM sleep behaviour disorder: A multicentre study. Brain. (2019) ; 142: : 744–759. |
10. | Berg D , Borghammer P , Fereshtehnejad SM , et al. Prodromal Parkinson disease subtypes – key to understanding heterogeneity. Nat Rev Neurol. (2021) ; 17: : 349–361. |
11. | Pilotto A , Romagnolo A , Tuazon JA , et al. Orthostatic hypotension and REM sleep behaviour disorder: Impact on clinical outcomes in α-synucleinopathies. J Neurol Neurosurg Psychiatry. (2019) ; 90: : 1257–1263. |
12. | Berg D and Postuma RB. From prodromal to overt Parkinson’s disease: Towards a new definition in the year 2040.J Parkinsons Dis (2018) ; 8: : S19–S23. |
13. | Donaghy PC , Carrarini C , Ferreira D , et al. Research diagnostic criteria for mild cognitive impairment with Lewy bodies: A systematic review and meta-analysis. Alzheimers Dement. (2023) ; 19: : 3186–3202. |
14. | Šubert M , Novotný M , Tykalová T , et al. Spoken language alterations can predict phenoconversion in isolated rapid eye movement sleep behavior disorder: A multicentric study. Ann Neurol. (2024) ; 95: : 530–543. |
15. | Vijiaratnam N and Foltynie T. How should we be using biomarkers in trials of disease modification in Parkinson’s disease? Brain (2023) ; 146: : 4845–4869. |
16. | Siderowf A , Concha-Marambio L , Lafontant DE , et al. Assessment of heterogeneity among participants in the Parkinson’s Progression Markers Initiative cohort using α-synuclein seed amplification: A cross-sectional study. Lancet Neurol. (2023) ; 22: : 407–417. |
17. | Eusebi P , Giannandrea D , Biscetti L , et al. Diagnostic utility of cerebrospinal fluid α-synuclein in Parkinson’s disease: A systematic review and meta-analysis. Mov Disord. (2017) ; 32: : 1389–1400. |
18. | Zhou B , Wen M , Yu W-F , et al. The diagnostic and differential diagnosis utility of cerebrospinal fluid α-synuclein levels in Parkinson’s disease: A meta-analysis. Parkinsons Dis. (2015) ; 2015: : 567386. |
19. | Sako W , Murakami N , Izumi Y , et al. Reduced alpha-synuclein in cerebrospinal fluid in synucleinopathies: Evidence from a meta-analysis. Mov Disord. (2014) ; 29: : 1599–1605. |
20. | Mollenhauer B , Parnetti L , Rektorova I , et al. Biological confounders for the values of cerebrospinal fluid proteins in Parkinson’s disease and related disorders. J Neurochem. (2016) ; 139 Suppl 1: : 290–317. |
21. | Parnetti L , Cicognola C , Eusebi P , et al. Value of cerebrospinal fluid α-synuclein species as biomarker in Parkinson’s diagnosis and prognosis. Biomark Med. (2016) ; 10: : 35–49. |
22. | Majbour NK , Vaikath NN , van Dijk KD , et al. Oligomeric and phosphorylated alpha-synuclein as potential CSF biomarkers for Parkinson’s disease. Mol Neurodegener. (2016) ; 11: : 7. |
23. | Maetzler W , Pilotto A , Apel A , et al. In vivo markers of Parkinson’s disease and dementia with Lewy bodies: Current value of the 5G4 α-synuclein antibody. Acta Neuropathol. (2014) ; 128: : 893–895. |
24. | Parnetti L , Gaetani L , Eusebi P , et al. CSF and blood biomarkers for Parkinson’s disease. Lancet Neurol. (2019) ; 18: : 573–586. |
25. | Andersen AD , Binzer M , Stenager E , et al. Cerebrospinal fluid biomarkers for Parkinson’s disease – a systematic review. Acta Neurol Scand. (2017) ; 135: : 34–56. |
26. | Aasly JO , Johansen KK , Brønstad G , et al. Elevated levels of cerebrospinal fluid α-synuclein oligomers in healthy asymptomatic LRRK2 mutation carriers. Front Aging Neurosci (2014) ; 6: : 248. |
27. | Majbour NK , Aasly JO , Hustad E , et al. CSF total and oligomeric α-Synuclein along with TNF-α as risk biomarkers for Parkinson’s disease: A study in LRRK2 mutation carriers. Transl Neurodegener. (2020) ; 9: : 15. |
28. | Wang X-T , Yu H , Liu F-T , et al. Associations of sleep disorders with cerebrospinal fluid α-synuclein in prodromal and early Parkinson’s disease. J Neurol. (2022) ; 269: : 2469–2478. |
29. | Zetterberg H . Neurofilament light: A dynamic cross-disease fluid biomarker for neurodegeneration. Neuron. (2016) ; 91: : 1–3. |
30. | Aamodt WW , Waligorska T , Shen J , et al. Neurofilament light chain as a biomarker for cognitive decline in Parkinson disease. Mov Disord. (2021) ; 36: : 2945–2950. |
31. | Choe CU , Niemann L , Englisch C , et al. Subclinical cardiac microdamage, motor severity, and cognition in Parkinson’s disease. Mov Disord. (2020) ; 35: : 1863–1868. |
32. | Ma LZ , Zhang C , Wang H , et al. Serum neurofilament dynamics predicts cognitive progression in de novo Parkinson’s disease. J Parkinsons Dis. (2021) ; 11: : 1117–1127. |
33. | Niemann L , Lezius S , Maceski A , et al. Serum neurofilament is associated with motor function, cognitive decline and subclinical cardiac damage in advanced Parkinson’s disease (MARK-PD). Parkinsonism Relat Disord. (2021) ; 90: : 44–48. |
34. | Lin CH , Li CH , Yang KC , et al. Blood NfL: A biomarker for disease severity and progression in Parkinson disease. Neurology. (2019) ; 93: : e1104–e1111. |
35. | Mollenhauer B , Dakna M , Kruse N , et al. Validation of serum neurofilament light chain as a biomarker of Parkinson’s disease progression. Mov Disord. (2020) ; 35: : 1999–2008. |
36. | Oosterveld LP , Verberk IMW , Majbour NK , et al. CSF or serum neurofilament light added to α-Synuclein panel discriminates Parkinson’s from controls. Mov Disord. (2020) ; 35: : 288–295. |
37. | Höglinger GU , Adler CH , Berg D , et al. A biological classification of Parkinson’s disease: The SynNeurGe research diagnostic criteria. Lancet Neurol. (2024) ; 23: : 191–204. |
38. | Hansson O , Janelidze S , Hall S , et al. Blood-based NfL: A biomarker for differential diagnosis of parkinsonian disorder. Neurology. (2017) ; 88: : 930–937. |
39. | Marques TM , van Rumund A , Oeckl P , et al. Serum NFL discriminates Parkinson disease from atypical parkinsonisms. Neurology. (2019) ; 92: : e1479–e1486. |
40. | Chelban V , Nikram E , Perez-Soriano A , et al. Neurofilament light levels predict clinical progression and death in multiple system atrophy. Brain. (2022) ; 145: : 4398–4408. |
41. | Nicklas WJ , Saporito M , Basma A , et al. Mitochondrial mechanisms of neurotoxicity. Ann N Y Acad Sci. (1992) ; 648: : 28–36. |
42. | Herbert MK , Eeftens JM , Aerts MB , et al. CSF levels of DJ-1 and tau distinguish MSA patients from PD patients and controls. Parkinsonism Relat Disord. (2014) ; 20: : 112–115. |
43. | Waragai M , Wei J , Fujita M , et al. Increased level of DJ-1 in the cerebrospinal fluids of sporadic Parkinson’s disease. Biochem Biophys Res Commun. (2006) ; 345: : 967–972. |
44. | Moors T , Paciotti S , Chiasserini D , et al. Lysosomal dysfunction and α-synuclein aggregation in Parkinson’s disease: Diagnostic links. Mov Disord. (2016) ; 31: : 791–801. |
45. | Nalls MA , Duran R , Lopez G , et al. A multicenter study of glucocerebrosidase mutations in dementia with Lewy bodies. JAMA Neurol. (2013) ; 70: : 727–735. |
46. | Lerche S , Schulte C , Wurster I , et al. The mutation matters: CSF profiles of GCase, sphingolipids, α-synuclein in PD(GBA). Mov Disord. (2021) ; 36: : 1216–1228. |
47. | Parnetti L , Chiasserini D , Persichetti E , et al. Cerebrospinal fluid lysosomal enzymes and alpha-synuclein in Parkinson’s disease. Mov Disord. (2014) ; 29: : 1019–1027. |
48. | Omer N , Giladi N , Gurevich T , et al. Glucocerebrosidase activity is not associated with Parkinson’s disease risk or severity. Mov Disord. (2022) ; 37: : 651–652. |
49. | Paslawski W , Khosousi S , Hertz E , et al. Large-scale proximity extension assay reveals CSF midkine and DOPA decarboxylase as supportive diagnostic biomarkers for Parkinson’s disease. Transl Neurodegener. (2023) ; 12: : 42. |
50. | Pereira JB , Kumar A , Hall S , et al. DOPA decarboxylase is an emerging biomarker for Parkinsonian disorders including preclinical Lewy body disease. Nat Aging. (2023) ; 3: : 1201–1209. |
51. | Verbeek MM and Bloem BR. An emerging biomarker for dopaminergic cell loss. Nat Aging. (2023) ; 3: : 1180–1182. |
52. | Mosley RL , Hutter-Saunders JA , Stone DK , et al. Inflammation and adaptive immunity in Parkinson’s disease. Cold Spring Harb Perspect Med. (2012) ; 2: : a009381. |
53. | Muñoz-Delgado L , Macías-García D , Jesús S , et al. Peripheral immune profile and neutrophil-to-lymphocyte ratio in Parkinson’s disease. Mov Disord. (2021) ; 36: : 2426–2430. |
54. | Olsson B , Constantinescu R , Holmberg B , et al. The glial marker YKL-40 is decreased in synucleinopathies. Mov Disord. (2013) ; 28: : 1882–1885. |
55. | Wennström M , Surova Y , Hall S , et al. The inflammatory marker YKL-40 is elevated in cerebrospinal fluid from patients with Alzheimer’s but not Parkinson’s disease or dementia with Lewy bodies. PLoS One. (2015) ; 10: : e0135458. |
56. | Abdo WF , van de Warrenburg BP , Kremer HP , et al. CSF biomarker profiles do not differentiate between the cerebellar and parkinsonian phenotypes of multiple system atrophy. Parkinsonism Relat Disord. (2007) ; 13: : 480–482. |
57. | Santaella A , Kuiperij HB , van Rumund A , et al. Cerebrospinal fluid myelin basic protein is elevated in multiple system atrophy. Parkinsonism Relat Disord. (2020) ; 76: : 80–84. |
58. | Prusiner SB , Woerman AL , Mordes DA , et al. Evidence for α-synuclein prions causing multiple system atrophy in humans with parkinsonism. Proc Natl Acad Sci U S A. (2015) ; 112: : E5308–5317. |
59. | Atarashi R , Sano K , Satoh K , et al. Real-time quaking-induced conversion: A highly sensitive assay for prion detection. Prion. (2011) ; 5: : 150–153. |
60. | Moda F , Gambetti P , Notari S , et al. Prions in the urine of patients with variant Creutzfeldt-Jakob disease. N Engl J Med. (2014) ; 371: : 530–539. |
61. | Concha-Marambio L , Pritzkow S , Shahnawaz M , et al. Seed amplification assay for the detection of pathologic alpha-synuclein aggregates in cerebrospinal fluid. Nat Protoc. (2023) ; 18: : 1179–1196. |
62. | Fontana E , Bongianni M , Benussi A , et al. Detection of TDP-43 seeding activity in the olfactory mucosa from patients with frontotemporal dementia. Alzheimers Dement. (2024) ; 20: : 1156–1165. |
63. | Kraus A , Saijo E , Metrick MA 2nd , et al. Seeding selectivity and ultrasensitive detection of tau aggregate conformers of Alzheimer disease. Acta Neuropathol. (2019) ; 137: : 585–598. |
64. | Grossauer A , Hemicker G , Krismer F , et al. α-synuclein seed amplification assays in the diagnosis of synucleinopathies using cerebrospinal fluid-a systematic review and meta-analysis. Mov Disord Clin Pract. (2023) ; 10: : 737–747. |
65. | Rossi M , Candelise N , Baiardi S , et al. Ultrasensitive RT-QuIC assay with high sensitivity and specificity for Lewy body-associated synucleinopathies. Acta Neuropathol. (2020) ; 140: : 49–62. |
66. | Brockmann K , Quadalti C , Lerche S , et al. Association between CSF alpha-synuclein seeding activity and genetic status in Parkinson’s disease and dementia with Lewy bodies. Acta Neuropathol Commun. (2021) ; 9: : 175. |
67. | Peña-Bautista C , Kumar R , Baquero M , et al. Misfolded alpha-synuclein detection by RT-QuIC in dementia with lewy bodies: A systematic review and meta-analysis. Front Mol Biosci. (2023) ; 10: : 1193458. |
68. | Pilotto A , Bongianni M , Tirloni C , et al. CSF alpha-synuclein aggregates by seed amplification and clinical presentation of AD. Alzheimers Dement. (2023) ; 19: : 3754–3759. |
69. | Singer W , Schmeichel AM , Shahnawaz M , et al. Alpha-synuclein oligomers and neurofilament light chain in spinal fluid differentiate multiple system atrophy from Lewy body synucleinopathies. Ann Neurol. (2020) ; 88: : 503–512. |
70. | Shahnawaz M , Mukherjee A , Pritzkow S , et al. Discriminating α-synuclein strains in Parkinson’s disease and multiple system atrophy. Nature. (2020) ; 578: : 273–277. |
71. | Quadalti C , Calandra-Buonaura G , Baiardi S , et al. Neurofilament light chain and α-synuclein RT-QuIC as differential diagnostic biomarkers in parkinsonisms and related syndromes. NPJ Parkinsons Dis. (2021) ; 7: : 93. |
72. | Iranzo A , Fairfoul G , Ayudhaya ACN , et al. Detection of α-synuclein in CSF by RT-QuIC in patients with isolated rapid-eye-movement sleep behaviour disorder: A longitudinal observational study. Lancet Neurol. (2021) ; 20: : 203–212. |
73. | Singer W , Schmeichel AM , Shahnawaz M , et al. Alpha-synuclein oligomers and neurofilament light chain predict phenoconversion of pure autonomic failure. Ann Neurol. (2021) ; 89: : 1212–1220. |
74. | Bongianni M , Catalan M , Perra D , et al. Olfactory swab sampling optimization for α-synuclein aggregate detection in patients with Parkinson’s disease. Transl Neurodegener. (2022) ; 11: : 37. |
75. | Orrù CD , Ma TC , Hughson AG , et al. A rapid α-synuclein seed assay of Parkinson’s disease CSF panel shows high diagnostic accuracy. Ann Clin Transl Neurol. (2021) ; 8: : 374–384. |
76. | Arnold SE , Lee EB , Moberg PJ , et al. Olfactory epithelium amyloid-beta and paired helical filament-tau pathology in Alzheimer disease. Ann Neurol. (2010) ; 67: : 462–469. |
77. | Witt M , Bormann K , Gudziol V , et al. Biopsies of olfactory epithelium in patients with Parkinson’s disease. Mov Disord. (2009) ; 24: : 906–914. |
78. | Brozzetti L , Sacchetto L , Cecchini MP , et al. Neurode-generation-associated proteins in human olfactory neurons collected by nasal brushing. Front Neurosci. (2020) ; 14: : 145. |
79. | Stefani A , Iranzo A , Holzknecht E , et al. Alpha-synuclein seeds in olfactory mucosa of patients with isolated REM sleep behaviour disorder. Brain. (2021) ; 144: : 1118–1126. |
80. | De Luca CMG , Elia AE , Portaleone SM , et al. Efficient RT-QuIC seeding activity for α-synuclein in olfactory mucosa samples of patients with Parkinson’s disease and multiple system atrophy. Transl Neurodegener. (2019) ; 8: : 24. |
81. | Bargar C , De Luca CMG , Devigili G , et al. Discrimination of MSA-P and MSA-C by RT-QuIC analysis of olfactory mucosa: The first assessment of assay reproducibility between two specialized laboratories. Mol Neurodegener. (2021) ; 16: : 82. |
82. | Beach TG , Adler CH , Sue LI , et al. Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. (2010) ; 119: : 689–702. |
83. | Sprenger FS , Stefanova N , Gelpi E , et al. Enteric nervous system α-synuclein immunoreactivity in idiopathic REM sleep behavior disorder. Neurology. (2015) ; 85: : 1761–1768. |
84. | Vilas D , Iranzo A , Tolosa E , et al. Assessment of α-synuclein in submandibular glands of patients with idiopathic rapid-eye-movement sleep behaviour disorder: A case-control study. Lancet Neurol. (2016) ; 15: : 708–718. |
85. | Iranzo A , Borrego S , Vilaseca I , et al. α-Synuclein aggregates in labial salivary glands of idiopathic rapid eye movement sleep behavior disorder. Sleep. (2018) ; 41: : doi: 10.1093/sleep/zsy101. |
86. | Doppler K , Jentschke HM , Schulmeyer L , et al. Dermal phospho-alpha-synuclein deposits confirm REM sleep behaviour disorder as prodromal Parkinson’s disease. Acta Neuropathol. (2017) ; 133: : 535–545. |
87. | Doppler K , Antelmi E , Kuzkina A , et al. Consistent skin α-synuclein positivity in REM sleep behavior disorder – A two center two-to-four-year follow-up study. Parkinsonism Relat Disord. (2021) ; 86: : 108–113. |
88. | Donadio V , Doppler K , Incensi A , et al. Abnormal α-synuclein deposits in skin nerves: Intra- and inter-laboratory reproducibility. Eur J Neurol. (2019) ; 26: : 1245–1251. |
89. | Al-Qassabi A , Tsao TS , Racolta A , et al. Immunohistochemical detection of synuclein pathology in skin in idiopathic rapid eye movement sleep behavior disorder and parkinsonism. Mov Disord. (2021) ; 36: : 895–904. |
90. | Antelmi E , Pizza F , Donadio V , et al. Biomarkers for REM sleep behavior disorder in idiopathic and narcoleptic patients. Ann Clin Transl Neurol. (2019) ; 6: : 1872–1876. |
91. | Donadio V , Incensi A , Rizzo G , et al. Skin biopsy may help to distinguish multiple system atrophy-parkinsonism from Parkinson’s disease with orthostatic hypotension. Mov Disord. (2020) ; 35: : 1649–1657. |
92. | Doppler K , Weis J , Karl K , et al. Distinctive distribution of phospho-alpha-synuclein in dermal nerves in multiple system atrophy. Mov Disord. (2015) ; 30: : 1688–1692. |
93. | Liguori R , Donadio V , Wang Z , et al. A comparative blind study between skin biopsy and seed amplification assay to disclose pathological α-synuclein in RBD. NPJ Parkinsons Dis. (2023) ; 9: : 34. |
94. | Kuzkina A , Panzer C , Seger A , et al. Dermal real-time quaking-induced conversion is a sensitive marker to confirm isolated rapid eye movement sleep behavior disorder as an early α-synucleinopathy. Mov Disord. (2023) ; 38: : 1077–1082. |
95. | Iranzo A , Mammana A , Muñoz-Lopetegi A , et al. Misfolded α-synuclein assessment in the skin and CSF by RT-QuIC in isolated REM sleep behavior disorder. Neurology. (2023) ; 100: : e1944–e1954. |
96. | Doppler K . Detection of dermal alpha-synuclein deposits as a biomarker for Parkinson’s disease. J Parkinsons Dis. (2021) ; 11: : 937–947. |
97. | Kuzkina A , Rößle J , Seger A , et al. Combining skin and olfactory α-synuclein seed amplification assays (SAA)-towards biomarker-driven phenotyping in synucleinopathies. NPJ Parkinsons Dis. (2023) ; 9: : 79. |
98. | Jiang C , Hopfner F , Berg D , et al. Validation of α-synuclein in L1CAM-immunocaptured exosomes as a biomarker for the stratification of parkinsonian syndromes. Mov Disord. (2021) ; 36: : 2663–2669. |
99. | Kluge A , Bunk J , Schaeffer E , et al. Detection of neuron-derived pathological α-synuclein in blood. Brain. (2022) ; 145: : 3058–3071. |
100. | Meloni M , Agliardi C , Guerini FR , et al. Oligomeric alpha-synuclein and STX-1A from neural-derived extracellular vesicles (NDEVs) as possible biomarkers of REM sleep behavior disorder in Parkinson’s disease: A preliminary cohort study. Int J Mol Sci. (2023) ; 24: : 8839. |
101. | Okuzumi A , Hatano T , Matsumoto G , et al. Propagative α-synuclein seeds as serum biomarkers for synucleinopathies. Nat Med. (2023) ; 29: : 1448–1455. |
102. | Schaeffer E , Kluge A , Schulte C , et al. Association of misfolded α-synuclein derived from neuronal exosomes in blood with Parkinson’s disease diagnosis and duration. J Parkinsons Dis. (2024) ; 14: : 667–679. |
103. | Kluge A , Schaeffer E , Bunk J , et al. Detecting misfolded α-synuclein in blood years before the diagnosis of Parkinson’s disease. Mov Disord. (2024) ; 39: : 1289–1299. |
104. | Ashton NJ , Janelidze S , Al Khleifat A , et al. A multicentre validation study of the diagnostic value of plasma neurofilament light. Nat Commun. (2021) ; 12: : 3400. |
105. | Bacioglu M , Maia LF , Preische O , et al. Neurofilament light chain in blood and CSF as marker of disease progression in mouse models and in neurodegenerative diseases. Neuron. (2016) ; 91: : 494–496. |
106. | Pilotto A , Imarisio A , Carrarini C , et al. Plasma neurofilament light chain predicts cognitive progression in prodromal and clinical dementia with Lewy bodies. J Alzheimers Dis. (2021) ; 82: : 913–919. |
107. | Galper J , Balwani M , Fahn S , et al. Cytokines and Gaucher biomarkers in glucocerebrosidase carriers with and without Parkinson disease. Mov Disord. (2021) ; 36: : 1451–1455. |
108. | Yan S , Jiang C , Janzen A , et al. Neuronally derived extracellular vesicle α-synuclein as a serum biomarker for individuals at risk of developing Parkinson disease. JAMA Neurol. (2024) ; 81: : 59–68. |
109. | Wilke C , Dos Santos MCT , Schulte C , et al. Intraindividual neurofilament dynamics in serum mark the conversion to sporadic Parkinson’s disease. Mov Disord. (2020) ; 35: : 1233–1238. |
110. | Zhang X , Ma L , Liang D , et al. Neurofilament light protein predicts disease progression in idiopathic REM sleep behavior disorder. J Parkinsons Dis. (2023) ; 13: : 485–499. |


