
Dystrophin Genotype and Risk of Neuropsychiatric Disorders in Dystrophinopathies: A Systematic Review and Meta-Analysis
Abstract
Background:
Dystrophinopathies are associated with neuropsychiatric disorders due to alterations in dystrophin/DMD expression.
Objective:
The objective was to estimate the association of developmental disorders, autism spectrum disorders (ASD), attention deficit hyperactivity disorder (ADHD), depression, anxiety disorders, and obsessive-compulsive disorder with the dystrophin/DMD genotype in population with dystrophinopathies.
Methods:
Systematic searches of Medline, Scopus, Web of Science, and Cochrane Library were performed from inception to September 2022. We included observational studies in the population with Becker or Duchenne muscular dystrophies (BMD, DMD) that estimated the prevalence of these disorders according to Dp140 and/or Dp71 genotype. Meta-analysis of the prevalence ratio (PR) of genotype comparisons was conducted for each disorder.
Results:
Ten studies were included in the systematic review. In BMD, Dp140+ vs. Dp140– and Dp71+ vs. Dp71– were associated with developmental disorders with a PR of 0.11 (0.04, 0.34) and 0.22 (0.07, 0.67), respectively. In DMD, Dp140+/Dp71+ vs. Dp140– /Dp71– had a PR of 0.40 (0.28, 0.57), and Dp71+ vs. Dp71– had a PR of 0.47 (0.36, 0.63) for ADHD. However, there was no association of genotype with ASD, only a trend was observed for Dp71+ vs. Dp71–, with a PR of 0.61 (0.35, 1.06). Moreover, the data showed no association of these isoforms with emotional-related disorders.
Conclusions:
In BMD, Dp140 and Dp71 could be associated with developmental disorders, while ADHD might be associated with the Dp71 genotype in DMD. Further research is needed regarding Dp140 and Dp71, especially in DMD for ASD.
INTRODUCTION
Dystrophinopathies are a group of diseases that include Becker muscular dystrophy (BMD) and Duchenne muscular dystrophy (DMD), caused by mutations in the dystrophin/DMD gene [1, 2], and have an incidence of one case per 18,500 male births and one per 3,500– 9,000 male births, respectively [3, 4]. Although both dystrophinopathies share some clinical characteristics, there are differences at the genetic level that largely determine the clinic and progression of each type of dystrophinopathy. In those with BMD, mutations do not affect the reading frame, and functional dystrophin is partially produced, while in those with DMD, mutations affect the reading frame, and there is no dystrophin. At the muscular level, alterations in dystrophin cause muscle necrosis and degeneration with replacement by fibrotic tissue, resulting in muscle contractures and decreased muscle strength, which are more pronounced in DMD. In the long term, there is early loss of ambulation and complications that limit life expectancy, including cardiac and respiratory failure [2, 5, 6]. Despite the improved use of glucocorticoids, new treatments, and possible genetic modifiers that could modulate the DMD phenotype, the prognosis in DMD remains poor [7– 9].
In addition to the above, patients with BMD/DMD have an increased risk of several cognitive and neuropsychiatric disorders [10]. This is due to alterations in the expression of full-length dystrophin (Dp427) and, depending on the mutation site, of two short isoforms expressed by internal promoters, Dp140 and Dp71, that have important roles in central nervous system (CNS) development, neuroplasticity and synaptogenesis. Thus, alterations in Dp427 cause dysfunctions in GABAergic transmission by reducing the clustering of postsynaptic GABAA receptors [10, 11]. It is suggested that Dp427 is preferentially expressed in the amygdala and in the CA4 region of the hippocampus [12, 13]. Alterations in Dp71 (promoter located 8 kb upstream of exon 63), which is ubiquitously expressed in the CNS, cause dysfunctions in postsynaptic glutaminergic receptors involved in glutaminergic transmission and alter the permeability of the blood– brain barrier [14]. Last, Dp140 (promoter between exons 44 and 45) is expressed during foetal development, with functions of transcription factor activity, neuronal differentiation and dendritic development [14].
A previous study from our center showed a prevalence of ≈6 and ≈20% of autism spectrum disorders (ASD) and attention deficit disorder and hyperactivity (ADHD) in dystrophinopathies [15]. Moreover, it is suggested that the number of affected isoforms is positively associated with the risk of neuropsychiatric disorders [10, 13]. Therefore, the purpose of this systematic review and meta-analysis was to determine the association of different genotypes and the prevalence of neurodevelopmental disorders, including ASD and ADHD, and emotional-related disorders, including depression, anxiety, obsessive-compulsive disorder (OCD) and emotional and behavioural dysregulation, among the population with BMD and DMD.
MATERIALS AND METHODS
This systematic review and meta-analysis was conducted following the Meta-analyses of Observational Studies in Epidemiology (MOOSE) guidelines [16] and the Cochrane Collaboration Handbook recommendations [17]. It was registered in PROSPERO (registration number: CRD42021284658).
Search strategy
A systematic search of the MEDLINE, Scopus, Web of Science, and Cochrane Library databases was conducted from inception until September 2022. We also searched the grey literature, i.e., Google Scholar, OpenGrey, Theseo, and the Networked Digital Library of Theses and Dissertations databases. Search terms included epidemiology, prevalence, survey, frequency, question*, ratio, rate, Becker muscular dystrophy, Duchenne muscular dystrophy, dystroph*, developmental disorders, neurodevelopmental disorders, attention deficit hyperactivity disorder, ADHD, autism spectrum disorder, autism, ASD, depression, anxiety, obsessive-compulsive disorder, obsessive compulsive disorder, OCD, mental disorders, and mental health. References of the included studies were reviewed to find additional studies. Study authors were contacted if necessary. The search was performed independently by two reviewers (C.P.-M. and I.C.-R.), and disagreements were resolved by consensus or with a third reviewer (V.M.-V.). The specific search is detailed in the supplementary appendix 1.
Inclusion/exclusion criteria
The inclusion criteria were as follows: i) participants: men with BMD or DMD, without age restrictions. Genotype must be available to form subgroups of participants according to mutation site in the DMD gene or expression of Dp427, Dp140, and Dp71; ii) exposure: Dp427– (all participants), Dp140+/Dp140–, Dp71+/Dp71–, mutations upstream of exon 44-45 (Dp140+), mutations downstream of exon 44-45 (Dp140–), mutations upstream of exon 63 (Dp71+), mutations downstream of exon 63 (Dp71–). Exceptionally, mutations upstream of exon 30 (Dp140+) and mutations between exons 45 and 55 (Dp140–) were considered even though they had interpretative limitations because they could improve genotype comparison; iii) design: observational studies, including prospective, retrospective, and cross-sectional studies; iv) outcome: prevalence of developmental disorders, ASD, ADHD, depression, anxiety, OCD, emotional and behavioural dysregulation by genotypes.
The exclusion criteria were as follows: i) cohorts of participants with BMD/DMD with exclusively neurological disorders; and ii) studies with a sample size of less than 30 so that the effect sizes/associations were not distorted.
The study selection was conducted independently by two reviewers (C.P.-M. and I.C.-R.), and disagreements were solved by consensus or by a third reviewer (V.M.-V.).
Data extraction
An ad hoc table was performed with the data extracted from the included studies: i) reference (author and year of publication); ii) country; iii) population (disease, sample and age), iv) genotypes (comparisons by affected isoforms); v) outcomes: prevalence ratio (PR) of developmental disorders, ASD, ADHD, depression, anxiety, OCD, and emotional and behavioural dysregulation.
The data extraction was performed independently by two reviewers (C.P.-M. and I.C.-R.).
Risk of bias assessment
We assessed the risk of bias using the Quality Assessment Tool for Observational Cohort and Cross-Sectional Studies from the United States National Institute of Health National Heart, Lung, and Blood Institute [18]. This tool has 14 items that include methodological and statistical issues and potential confounders. Each item receives a score of poor/high risk, fair/some concerns/not applicable, or good/low risk. Due to the nature of the studies, some items may not be assessable. The overall score was determined by the number of domains with risk of bias. If >2 domains indicated high risk, the overall score was poor; if 2 domains indicated high risk, the overall score was fair; and if <2 domains indicated high risk, the overall score was good.
Two reviewers (C.P.-M. and I.C.-R.) assessed the risk of bias. Disagreements were resolved by consensus or by a third reviewer (V.M.-V.).
Grading the quality of evidence
We assessed the quality of evidence with the Grading of Recommendations, Assessment, Development and Evaluation (GRADE) tool [19]. The GRADE tool rates the strength of evidence as high, moderate, low, or very low based on the study design, risk of bias, indirect evidence, imprecision, inconsistency, and other variables.
Data synthesis
An ad hoc table was performed in which each genotype’s and outcome’s results were summarised. PR was estimated from the disorder’s prevalence in a genotype divided by its prevalence in another genotype.
When two or more genotypes could be compared for the same outcome and population (i.e., BMD or DMD), a random effects meta-analysis was conducted [20, 21]. We assessed the heterogeneity with the I2 statistic. The heterogeneity was deemed not important if the I2 statistic was 0– 30%, moderate if the I2 statistic was 30– 50%, substantial if the I2 statistic was 50– 75%, and considerable if the I2 statistic was 75– 100% [17, 22]. The p value of heterogeneity was also assessed and was statistically significant if p < 0.05. Because of the lack of studies, we were unable to assess the small study effect, publication bias [23] or subgroup studies according to age.
The statistical analyses were conducted using Stata software, version 15 (StataCorp, College Station, TX, USA).
Secondary analyses
Where possible, meta-analyses of prevalence by genotype for primary outcomes were performed. A meta-analysis of PR including dystrophinopathies cohort studies without separating BMD and DMD was also performed.
Modifications to the initial protocol
Although the registered protocol initially included the outcomes ASD, ADHD, depression, anxiety disorders, and OCD, it was decided to broaden the search to other disorders, which included developmental disorders and emotional and behavioural dysregulation. The outcome “developmental disorders” includes several outcomes identified as “neurodevelopmental disorders” in the diagnostic and statistical manual of mental disorders (DSM-V). It was considered a posteriori as an outcome in its own right, since genotypes were not available for specific disorders (e.g., ASD), only for “developmental disorders”. This was due to the need to expand the literature and to better interpret the data obtained.
RESULTS
Of the 1,736 studies identified, 10 were included in the systematic review (Table 1, Fig. 1) [24– 33], 8 were included in the meta-analyses [25– 32], and 10 were excluded for the reasons specified in the supplementary table 1.
Fig. 1
PRSIMA flowchart of study selection.
Table 1
Baseline characteristics of the participants in the included studies
| Reference | Count | Population | Genotype | Outcome | ||||||||
| Dis | N | Age | DD | ASD | ADHD | DEP | ANX | OCD | EBD | |||
| Lambert JT et al. (2020) [24] | US | BMD | 70 | 13.4±6.8 | Dp140+/Dp71+ vs. Dp140– /Dp71+ | – | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Mori-Yoshimura M et al. (2018) [25] | Japan | BMD | 125 | 37.2 | Dp140+ vs. Dp140– | ✓ | – | – | – | – | – | – |
| Dp71+ vs. Dp71– | ||||||||||||
| Mori-Yoshimura M et al. (2019) [26] | Japan | BMD | 76 | 38.8±13.3 | Dp140+ vs. Dp140– | ✓ | – | – | – | – | – | – |
| Dp71+ vs. Dp71– | ||||||||||||
| Colombo P et al. (2017) [27] | Italy | DMD | 47 | 10.9±3.9 | Dp140+ vs. Dp140– | ✓ | – | – | – | – | – | |
| Darmahkasih AJ et al. (2019) [28] | US | DMD | 700 | 13.0±5.6 | Dp140+/Dp71+ vs. Dp140– /Dp71– | – | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| Dp71+ vs. Dp71– | ||||||||||||
| Pane M et al. (2012) [29] | Italy | DMD | 103 | 12.6 | Dp140+ vs. Dp140– | – | – | ✓ | – | – | – | – |
| Dp140+/Dp71+ vs. Dp140– /Dp71+ | ||||||||||||
| Dp140+/Dp71+ vs. Dp140– /Dp71– | ||||||||||||
| Dp71+ vs. Dp71– | ||||||||||||
| Ricotti V et al. (2015) [30] | UKItalyBelgium | DMD | 130 | 9.8 | Dp140+/Dp71+ vs. Dp140– /Dp71– | – | ✓ | – | – | – | – | – |
| Dp71+ vs. Dp71– | ||||||||||||
| Saito Y et al. (2022) [31] | Japan | DMD | 55 | 9.4 | Dp140+ vs. Dp140– | – | ✓ | ✓ | – | – | – | – |
| Dp140+/Dp71+ vs. Dp140– /Dp71– | ||||||||||||
| Dp71+ vs. Dp71– | ||||||||||||
| Thangarajh M et al. (2019) [32] | FOR-DMD | DMD | 196 | 5.8±1.0 | Dp140+ vs. Dp140– | – | ✓ | ✓ | – | – | – | – |
| Fujino H et al. (2018) [33] | Japan | BMD + DMD | 56 | 12.9±5.2 | Dp140+ vs. Dp140– | – | ✓ | – | – | – | – | – |
| Dp140– /Dp71+ vs. Dp140– /Dp71– | ||||||||||||
| Dp140+/Dp71+ vs. Dp140– /Dp71+ | ||||||||||||
| Dp140+/Dp71+ vs. Dp140– /Dp71– | ||||||||||||
| Dp71+ vs. Dp71– | ||||||||||||
Count: country; Dis: disease; US: United States; UK: United Kingdom; N: sample size; DD: developmental disorders; Depression; ANX: anxiety disorders; EBD: emotional and behavioural dysregulation.
Four studies included cohorts from Japan [25, 26, 31, 33], three studies from Italy [27, 29, 30], two studies from the US [24, 28], one study each from Belgium and the UK [30], and one study from the Finding the Optimal Regimen for Duchenne Muscular Dystrophy (FOR-DMD) trial cohort [32]. Four studies included the BMD population, and seven included the DMD population, adding 1,558 participants with a minimum mean age of 5.8 years and a maximum mean age of 38.8 years. Seven studies included ASD, five studies included ADHD, and two studies included developmental disorders, depression, anxiety disorders, OCD, and emotional and behavioural dysregulation for each outcome. As described in Supplementary Table 2, the diagnostic method for DMD and BMD was not always described in detail. However, all studies included participants with DMD or BMD confirmed by genetic testing and/or biopsy and/or clinical criteria. Data collection and diagnostic criteria for neuropsychiatric disorders are included in Supplementary Table 3 and Supplementary Table 4.
Systematic review
Table 2 summarises the results obtained for each outcome and genotype.
Table 2
Results of the prevalence ratio by genotype-outcome in each study
| Reference | Disease | Genotype | Prevalence ratio |
| (95% CI) | |||
| 1 – Developmental disorders | |||
| Mori-Yoshimura M et al. (2018) [25] | BMD | Dp140+ vs. Dp140– | 0.13 (0.04, 0.43) |
| Mori-Yoshimura M et al. (2019) [26] | BMD | Dp140+ vs. Dp140– | 0.05 (0.00, 0.81) |
| Mori-Yoshimura M et al. (2018) [25] | BMD | Dp71+ vs. Dp71– | 0.26 (0.06, 1.06) |
| Mori-Yoshimura M et al. (2019) [26] | BMD | Dp71+ vs. Dp71– | 0.15 (0.02, 1.05) |
| 2 – Autism spectrum disorders | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 1.31 (0.34, 5.00) |
| Colombo P et al. (2017) [27] | DMD | Dp140+ vs. Dp140– | 1.12 (0.28, 4.45) |
| Saito Y et al. (2022) [31] | DMD | Dp140+ vs. Dp140– | 1.08 (0.66, 1.76) |
| Thangarajh M et al. (2019) [32] | DMD | Dp140+ vs. Dp140– | 0.24 (0.01, 4.90) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 0.43 (0.15, 1.24) |
| Ricotti V et al. (2015) [30] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 0.36 (0.10, 1.24) |
| Saito Y et al. (2022) [31] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.14 (0.40, 3.26) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 0.49 (0.19, 1.26) |
| Ricotti V et al. (2015) [30] | DMD | Dp71+ vs. Dp71– | 0.44 (0.17, 1.15) |
| Saito Y et al. (2022) [31] | DMD | Dp71+ vs. Dp71– | 1.10 (0.40, 3.02) |
| Fujino H et al. (2018) [33] | BMD + DMD | Dp140– /Dp71+ vs. Dp140– /Dp71– | 2.12 (0.14, 32.51) |
| Fujino H et al. (2018) [33] | BMD + DMD | Dp140+ vs. Dp140– | 1.02 (0.28, 3.76) |
| Fujino H et al. (2018) [33] | BMD + DMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 0.88 (0.24, 3.21) |
| Fujino H et al. (2018) [33] | BMD + DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.94 (0.12, 31.74) |
| Fujino H et al. (2018) [33] | BMD + DMD | Dp71+ vs. Dp71– | 1.98 (0.13, 29.34) |
| 3 – Attention deficit disorder and hyperactivity | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 1.71 (0.93, 3.15) |
| Pane M et al. (2012) [29] | DMD | Dp140+ vs. Dp140– | 0.64 (0.31, 1.32) |
| Saito Y et al. (2022) [31] | DMD | Dp140+ vs. Dp140– | 1.62 (0.11, 24.53) |
| Thangarajh M et al. (2019) [33] | DMD | Dp140+ vs. Dp140– | 0.11 (0.01, 1.93) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 0.42 (0.28, 0.62) |
| Pane M et al. (2012) [29] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 0.32 (0.15, 0.70) |
| Saito Y et al. (2022) [31] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.14 (0.06, 20.18) |
| Pane M et al. (2012) [29] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 0.77 (0.36, 1.65) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 0.51 (0.37, 0.71) |
| Pane M et al. (2012) [29] | DMD | Dp71+ vs. Dp71– | 0.39 (0.23, 0.65) |
| Saito Y et al. (2022) [31] | DMD | Dp71+ vs. Dp71– | 0.48 (0.03, 8.69) |
| 4 – Depression | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 0.73 (0.08, 6.60) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.51 (0.50, 4.60) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 1.44 (0.49, 4.26) |
| 5 – Anxiety disorders | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 1.91 (0.79, 4.60) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.34 (0.64, 2.79) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 1.48 (0.73, 3.03) |
| 6 – Obsessive-compulsive disorder | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 0.60 (0.18, 1.92) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 1.31 (0.57, 3.00) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 1.36 (0.61, 3.04) |
| 7 – Emotional and behavioural dysregulation | |||
| Lambert JT et al. (2020) [24] | BMD | Dp140+/Dp71+ vs. Dp140– /Dp71+ | 1.09 (0.59, 2.03) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp140+/Dp71+ vs. Dp140– /Dp71– | 0.69 (0.44, 1.07) |
| Darmahkasih AJ et al. (2019) [28] | DMD | Dp71+ vs. Dp71– | 0.80 (0.53, 1.19) |
In BMD, Dp140+ vs. Dp140– was associated with developmental disorders, and Dp71+ vs. Dp71– tended to be associated. In DMD, only Dp71+ vs. Dp71– (with or without Dp140) tended to be associated with ASD prevalence [28, 30], while Dp140+ vs. Dp140– did not obtain association or tendency to association. Conversely, Dp140+ vs. Dp140– tended to be associated with ADHD in DMD, and Dp71+ vs. Dp71– genotypes (with or without Dp140) were significantly associated with a lower prevalence of ADHD in two studies [28, 29].
Regarding emotional disorders, the studied genotypes were not associated with depression, anxiety disorders or OCD, nor was there a clear trend suggesting an association.
Risk of bias assessment
According to the Quality Assessment Tool for Observational Cohort and Cross-Sectional Studies from the United States National Institute of Health National Heart, Lung, and Blood Institute, all studies were scored as “good” in overall bias. By domain, 3 of 10 studies (30.0%) were assessed as some concerns regarding the use of standardised diagnostic criteria. Moreover, the sufficient timeframe in which an association could reasonably be expected was deemed to be problematic in 1 of 10 studies (10.0%). Because of the nature of the studies, the sample size calculation was not justified, nor were the tests repeated more than once or the investigators blinded. The risk of bias assessment is detailed in the Supplementary Table 5.
Quality of evidence assessment
According to the GRADE tool, the certainty of quality was moderate for developmental disorders in BMD and Dp140+/Dp71+ vs. Dp140–/Dp71– and Dp71+ vs. Dp71– for ADHD in DMD. The rest of the outcomes obtained a low certainty of quality. The quality of evidence assessment is detailed in the Supplementary Table 6.
Meta-analysis
In BMD, Dp140+ vs. Dp140– and Dp71+ vs. Dp71– were associated with developmental disorders, with PR = 0.11 (0.04, 0.34) and PR = 0.22 (0.07, 0.67), respectively (Fig. 2a). In DMD, no statistically significant associations were found for ASD (Fig. 2b). However, Dp140+/Dp71+ vs. Dp140– /Dp71– and Dp71+ vs. Dp71– were associated with ADHD, with PR = 0.40 (0.28, 0.57) and PR = 0.47 (0.36, 0.63), respectively (Fig. 2c).
Fig. 2
Meta-analysis of the prevalence ratio and 95% confidence interval by genotype comparisons of developmental disorders in Becker muscular dystrophy (a), autism spectrum disorders in Duchenne muscular dystrophy (b), and attention deficit hyperactivity disorder in Duchenne muscular dystrophy (c).
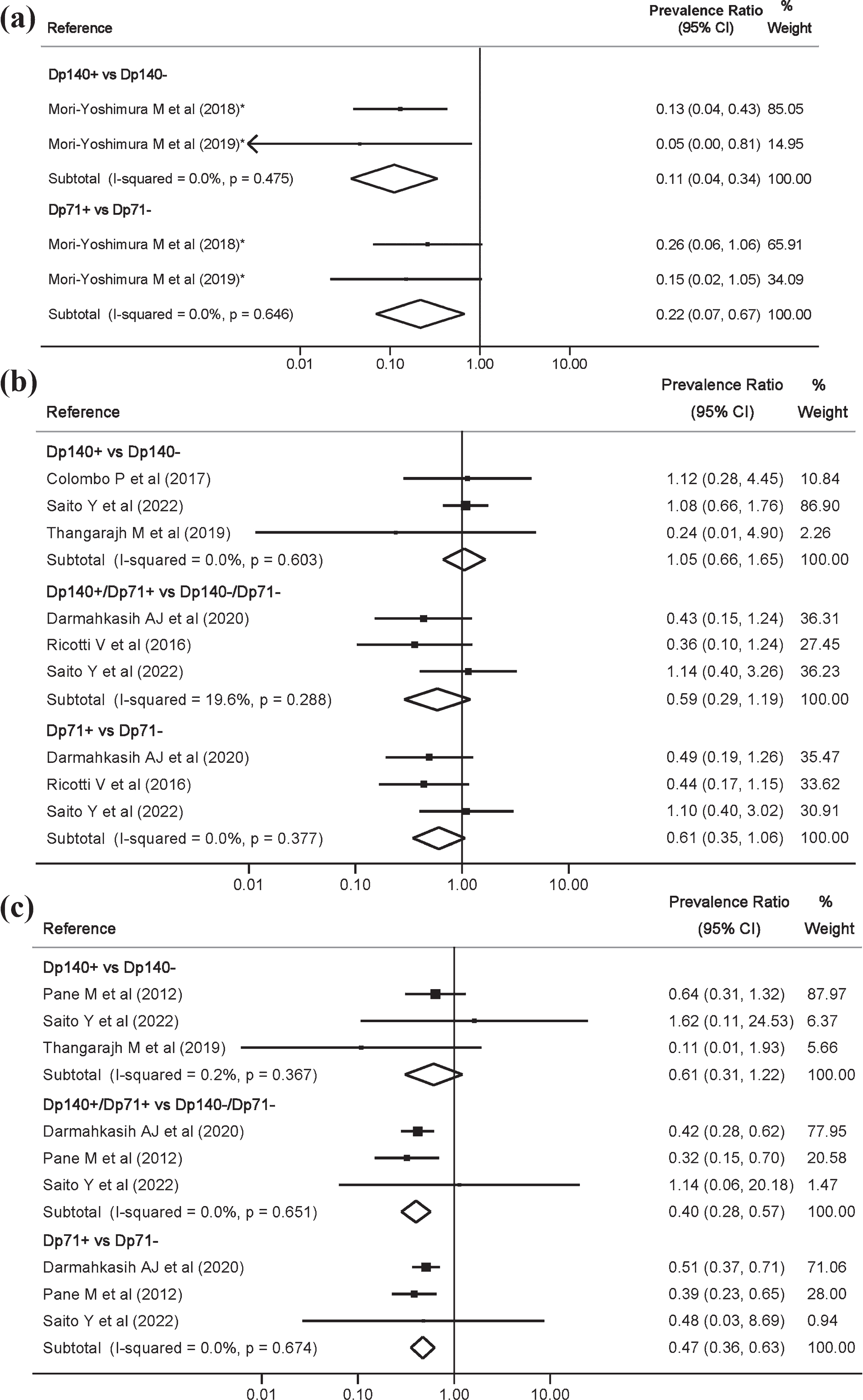
Heterogeneity for the main outcomes was not important and, overall, of 0%.
Secondary analyses
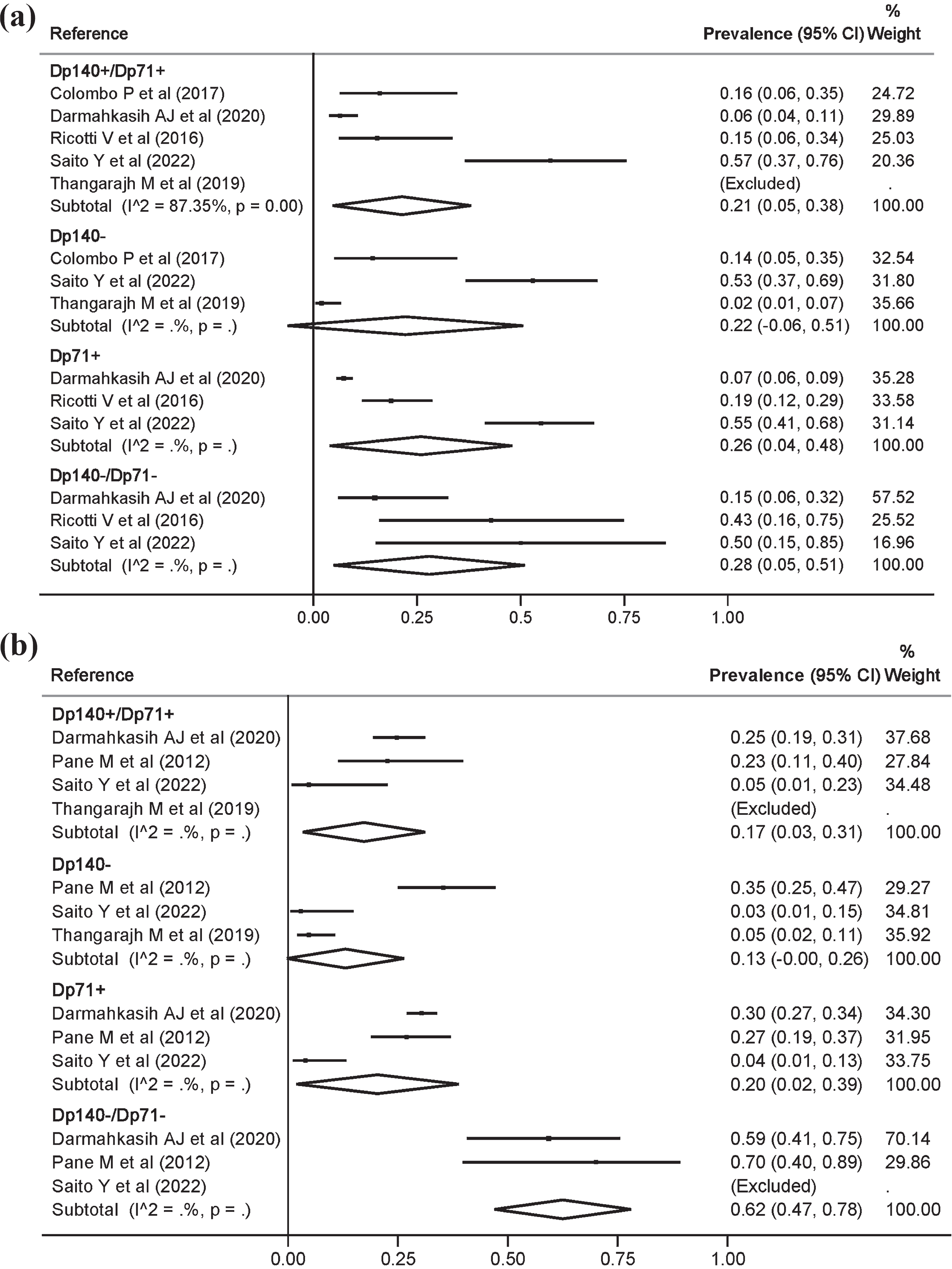
For ASD there were no statistically significant differences in prevalence by genotype (Fig. 3a). However, for ADHD a prevalence of 17.0% (3.0, 31.0) and 62.0% (47.0, 78.0) was estimated in the Dp140+/Dp71+ and Dp140– /Dp71– genotypes, respectively (Fig. 3b). Although heterogeneity could not be estimated for most genotype comparisons, visually some variability in estimates was observed. Regarding genotype comparisons with the inclusion of studies without considering the type of dystrophinopathy, the results were quite similar to those obtained in the main meta-analyses (Supplementary Figure 1).
Fig. 3
Meta-analysis of the prevalence and 95% confidence interval by genotype of autism spectrum disorders (a), and attention deficit hyperactivity disorder (b), in Duchenne muscular dystrophy.
DISCUSSION
Main findings
This systematic review and meta-analysis examined the association between genotype and the risk of neuropsychiatric disorders in BMD and DMD, including developmental disorders, ASD, ADHD, depression, anxiety disorders, OCD, and emotional and behavioural dysregulation. For developmental disorders, our data showed reductions of 89% and 78% when Dp140 and Dp71 were not altered in BMD, respectively. Conversely, in DMD, there was a reduction of 53% in the prevalence of ADHD for Dp71+ vs. Dp71–, a result that hardly varies when considering Dp140+/Dp71+ vs. Dp140– /Dp71–. Moreover, although it did not reach statistical significance, alterations in Dp71 also tended to be associated with ASD prevalence. Finally, Dp140 did not seem to have a relevant influence on the estimates.
Interpretation
Our meta-analysis suggests that the Dp71 isoform modulates the risk of developmental disorders in BMD and in DMD. In DMD, although the meta-analysis did not show a statistically significant association for ASD, a clear trend towards a deleterious association was observed when Dp71 was affected. Thus, more studies are needed to confirm or refute this possible association. However, Dp140 did not seem to have an influence in ASD. These results, and especially the null influence of Dp140, were replicated in the meta-analysis without considering the type of dystrophinopathy. This hypothesis would be consistent with the functional role of Dp71 in the CNS. Cell adhesion mediated by β1-integrin modulates synaptogenesis and neuronal migration during CNS development [34]. Dp71 modulates cell migration by binding to focal adhesion kinase and β1-integrin. In fact, in the absence of Dp71, fibroblast cell lines migrate at a faster rate [35]. The faster migration rate is supported by brain heterotopia, with abnormal dendritic development and impaired neuronal migration observed in DMD, which are characteristic changes that also occur in autism [36, 37].
Participants with both isoforms affected also had an increased risk of ADHD in DMD, with Dp71 being the isoform that reached statistical significance. Moreover, although some influence of Dp140 cannot be ruled out [29], the secondary meta-analysis in dystrophinopathies showed no association of Dp140, which requires further research, especially in relation to the possible role of Dp140 in ADHD in DMD, where it showed a tendency for a deleterious association when Dp140 was altered. These findings are supported by the functions of dystrophin in the CNS. In DMD, frontal and cerebellar white matter are altered, and cortical thickness is reduced. Both effects are typical in ADHD. Furthermore, the absence of Dp427 and Dp71 could affect dopaminergic cells in the ventral, tegmental, and pars compacta areas that receive GABAergic and glutaminergic inputs. Furthermore, alterations in the function of D4 receptors (which are expressed in the precerebral cortex) may be associated with ADHD. Interestingly, D4 receptor function may be involved in glutamate receptor-dependent plasticity [38, 39], and the latter could be associated with Dp71. Conversely, it is suggested that the lack of Dp427p and Dp140 in the cortex and cerebellum could cause cerebellar dysfunctions, which could increase the risk of ADHD and ASD [13, 40].
Regarding emotional-related disorders, our findings do not suggest an association between the isoforms affected and the risk of prevalence of emotional and behavioural dysregulation, depression, anxiety disorders, and OCD, although some association cannot be ruled out for the first outcome in DMD. There are several reasons that could explain these findings. First, it could be that the isoform most influential in these disorders is Dp427. Lack of Dp427 alters neuronal excitability and behaviour by altering the GABAA receptor clusters in the amygdala and hippocampus [41]. Moreover, it could affect dysbindin function, the dysfunction of which is associated with depression and psychosis [42]. Some authors have also considered that this type of disorder could develop partly because of disease awareness that is acquired over time [43], and in the case of Dp71– and its association with cognitive problems, disease awareness could be limited. Conversely, living with the disease itself may be a decisive factor in the development of mood disorders. Moreover, some disorders, such as OCD, appear later than others such as ASD and ADHD. Finally, young children may develop emotional disorders that manifest as oppositional behaviours or anger outbursts rather than the classic clinical presentation of depression or anxiety.
The results of this study have two relevant implications. First, contrary to what occurs with intellectual disability, in which an association with the number of dystrophin isoforms affected has been observed, in neuropsychiatric disorders this association is not so clear. Therefore, patient genotyping cannot currently be used to determine who should be evaluated more frequently or thoroughly, and international guidelines should be followed [10]. Second, some authors have suggested that some typical ASD behaviours in DMD improve with age [31]. This would be interesting to confirm with future longitudinal studies since, regardless of the possible influence of the genotype, it would provide greater knowledge of the disease and its progression and, ultimately, lead to better management of these patients.
Finally, it is worth highlighting an aspect of our study that should be considered for future research. Our study was an association study, and establishing causality from an association is complicated. Given that participants are born with a genotype that precedes pathology, probable causality can be established, supported by some pathophysiological mechanisms. However, our estimates are not deterministic, only probabilistic, and more research is needed in this regard, i.e., although a genotype may be associated with pathology, e.g., Dp71– with ADHD in DMD, having the Dp71– genotype is not a unique condition for having ADHD, nor does having Dp71+ exclude a higher risk of ADHD than in the rest of the population. Therefore, future research would be necessary, first, to increase the available evidence on the effect of Dp140 and Dp71 in neuropsychiatric disorders, since the available evidence is still limited, and second, to find other factors associated with these disorders, both at the genetic level (e.g., other genes, epigenetics, etc.), and at the histological and structural level of the CNS that explain why some people with DMD develop a certain disorder and others do not.
Limitations
Some limitations should be acknowledged. First, the scarcity of studies limits the statistical power of the meta-analyses and the interpretation of the systematic review. In the meta-analysis of ADHD, there is an overrepresentation of a study [28] in the pooled, due to the large sample size. Second, there is variability in the participants’ diagnostic methods for dystrophinopathy. All were diagnosed according to the usual guidelines (genetic testing and/or muscle biopsy, although biopsy is currently in disuse because it is an invasive technique), but it cannot be ruled out that any participant with DMD has a diagnosis of BMD, or vice versa (i.e., the reading frame rule is not always met). Moreover, with muscle biopsy, and especially with Western Blot in DMD, the amount of dystrophin (if there is any expression) correlates with the phenotype, but in BMD this correlation is more doubtful. Finally, there are cases of confirmed BMD with a severe phenotype, showing a diffuse spectrum in the differentiation of DMD and BMD. Third, different diagnostic criteria or data collection methods were used in each study, including self-reported questionnaires of the disorders suffered by the participants, which may eventually limit the interpretability of estimates [25]. Furthermore, the results of “developmental disorders” in the BMD are difficult to interpret, as it includes several outcomes (intellectual disability, ASD, etc.) that could not be analysed separately. Fourth, the difference in mean age between studies might distort the prevalence of disorders according to genotype. Fifth, the assessment of publication bias could not be performed. Therefore, despite the consistency of the results obtained and the mechanisms that would explain them, their existence cannot be completely ruled out. Sixth, cognitive problems should also be considered, which, due to their association with the affected isoforms, could slightly modify the final estimates. Seventh, the results could not be controlled by the use of glucocorticoids. However, previous research suggests that although corticosteroids can alter mood, this effect seems to become stabilised after a long period of time [44]. Eighth, to increase the number of comparisons, participants with mutations between exons 45 and 55 were considered to be Dp140–, as is widely accepted. However, this is controversial, since mutations between exons 45 and 51 could, on certain occasions, not predict a loss of Dp140 [45].
In conclusion, our study shows that ADHD are twice as prevalent in participants with DMD without the Dp71 isoform. The results also suggested, perhaps, an influence of Dp71 in ASD, but future studies are needed to confirm or refute this. Moreover, in BMD, the evidence also suggests that Dp71, and probably Dp140, are associated with developmental disorders. In contrast, no association with emotional disorders was found, requiring future research to clarify the role of dystrophin isoforms in these disorders or whether the development of these disorders is due to psychosocial factors derived from suffering from this type of disease. Dp140 was also not associated with neuropsychiatric disorders in DMD, although the scarcity of studies limits the validity of this statement. Therefore, further research is needed, especially for ADHD.
STATEMENTS AND DECLARATIONS
Contributions
Conceptualisation: C.P.-M.; methodology: C.P-M. and I.C.-R.; data curation and investigation: C.P.-M. and I.C.-R.; formal analysis: C.P.-M., E.J.-L., I.S.-D., and J.F.-B.-R.; validation and visualisation: E.J.-L., I.S.-D., and J.F.-B.-R.; writing— original draft preparation: C.P.-M., I.C.-R., and V.M.-V.; writing— review and editing: all authors; supervision: I.C.-R. and V.M.-V.; funding acquisition: V.M.-V.; project administration: V.M.-V. All authors have read and agreed to the published version of the manuscript.
Funding
C.P.-M. is supported by a grant from the Universidad de Castilla-La Mancha (2018-CPUCLM-7939) and by the European Regional Development Fund.
Conflicts of interest
The authors declare no conflicts of interest, and no funding was received for this study.
Data statement
The original contributions presented in the study are included in the article and supplementary material. Further inquiries can be directed to the corresponding author/s.
Acknowledgments
Not applicable.
Ethical approval
Not applicable.
SUPPLEMENTARY MATERIAL
[1] The supplementary material is available in the electronic version of this article: https://dx.doi.org/10.3233/JND-221586.
REFERENCES
[1] | Holland A , Carberry S , Ohlendieck K . Proteomics of the Dystrophin-glycoprotein Complex and Dystrophinopathy. Curr Protein Pept Sci [Internet]. (2014) [cited 2021 Apr 17];14: (8):680–97. Available from: https://pubmed.ncbi.nlm.nih.gov/24106963/. |
[2] | Waldrop MA , Flanigan KM . Update in Duchenne and Becker muscular dystrophy. Curr Opin Neurol [Internet]. (2019) [cited 2022 Mar 27];32: (5):722–7. Available from: https://pubmed.ncbi.nlm.nih.gov/31343429/. |
[3] | Bushby KM , Thambyayah M , Gardner-Medwin D . Prevalence and incidence of Becker muscular dystrophy. Lancet (London, England). (1991) ;337: (8748):1022–4. |
[4] | Mah JK , Korngut L , Dykeman J , Day L , Pringsheim T , Jette N . A systematic review and meta-analysis on the epidemiology of Duchenne and Becker muscular dystrophy. Neuromuscul Disord [Internet]. (2014) [cited 2020 Mar 26];24: (6):482–91. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24780148. |
[5] | Birnkrant DJ , Bushby K , Bann CM , Apkon SD , Blackwell A , Brumbaugh D , et al. Diagnosis and management of Duchenne muscular dystrophy, part Diagnosis, and neuromuscular, rehabilitation, endocrine, and gastrointestinal and nutritional management. Vol. 17, The Lancet Neurology. Lancet Publishing Group; (2018) , pp. 251–67. |
[6] | Birnkrant DJ , Bushby K , Bann CM , Alman BA , Apkon SD , Blackwell A , et al. Diagnosis and management of Duchenne muscular dystrophy, part Respiratory, cardiac, bone health, and orthopaedic management. Lancet Neurol [Internet]. (2018) [cited 2019 May 13];17: (4):347–61. Available from: http://www.ncbi.nlm.nih.gov/pubmed/29395990. |
[7] | Pascual-Morena C , Cavero-Redondo C , Saz-Lara A , Sequí-Domínguez I , Lucerón-Lucas-Torres M , Martínez-Vizcaíno V . Genetic modifiers and phenotype of duchenne muscular dystrophy: A systematic review and meta-analysis. Pharmaceuticals (Basel) [Internet]. (2021) [cited 2021 Nov 2];14: (8). Available from: https://pubmed.ncbi.nlm.nih.gov/34451895/. |
[8] | Goemans N . How glucocorticoids change life in Duchenne muscular dystrophy [Internet]. The Lancet. Lancet Publishing Group. (2018) [cited 2021 Apr 17];391: :406–7. Available from: https://pubmed.ncbi.nlm.nih.gov/29174483/. |
[9] | Pascual-Morena C , Cavero-Redondo I , Álvarez-Bueno C , Mesas AE , Pozuelo-Carrascosa D , Martínez-Vizcaíno V . Restorative treatments of dystrophin expression in Duchenne muscular dystrophy: A systematic review. Ann Clin Transl Neurol [Internet]. (2020) [cited 2020 Sep 19]; 7: (9):acn3.51149. Available from:https://onlinelibrary.wiley.com/doi/abs/10.1002/acn3.51149. |
[10] | Birnkrant DJ , Bushby K , Bann CM , Apkon SD , Blackwell A , Colvin MK , et al. Diagnosis and management of Duchenne muscular dystrophy, part Primary care, emergency management, psychosocial care, and transitions of care across the lifespan. LANCET Neurol. (2018) ;17: (5):445–55. |
[11] | Sekiguchi M , Zushida K , Yoshida M , Maekawa M , Kamichi S , Yoshida M , et al. A deficit of brain dystrophin impairs specific amygdala GABAergic transmission and enhances defensive behaviour in mice. Brain. (2009) ;132: (Pt 1):124–35. |
[12] | Doorenweerd N . Combining genetics, neuropsychology and neuroimaging to improve understanding of brain involvement in Duchenne muscular dystrophy - a narrative review. Neuromuscul Disord [Internet]. (2020) ;30: (6):437–42. Available from: https://pubmed.ncbi.nlm.nih.gov/32522501/. |
[13] | Doorenweerd N , Mahfouz A , van Putten M , Kaliyaperumal R , T’ Hoen PAC , Hendriksen JGM , et al. Timing and localization of human dystrophin isoform expression provide insights into the cognitive phenotype of Duchenne muscular dystrophy. Sci Rep. (2017) ;7: (1):12575. |
[14] | Daoud F , Candelario-Martínez A , Billard J-M , Avital A , Khelfaoui M , Rozenvald Y , et al. Role of mentalretardation-associated dystrophin-gene product Dp71 in excitatory synapse organization, synaptic plasticity and behavioral functions. PLoS One. (2009) ;4: (8):e6574. |
[15] | Pascual-Morena C , Cavero-Redondo I , Reina-Gutiérrez S , Saz-Lara A , López-Gil JF , Martínez-Vizcaíno V . Prevalence of neuropsychiatric disorders in duchenne and becker musculardystrophies: A systematic review and meta-analysis. Arch Phys Med Rehabil. 2022. |
[16] | Stroup DF , Berlin JA , Morton SC , Olkin I , Williamson GD , Rennie D , et al. Meta-analysis of observational studies in epidemiology: A proposal for reporting. J Am Med Assoc [Internet]. (2000) [cited 2021 Apr 17];283: (15):2008–12. Available from: https://pubmed.ncbi.nlm.nih.gov/10789670/. |
[17] | Higgins JP , Green S . Cochrane Handbook for Systematic Reviews of Interventions: Cochrane Book Series [Internet]. Cochrane Handbook for Systematic Reviews of Interventions: Cochrane Book Series. John Wiley and Sons; (2008) [cited 2020 Oct 28]; pp. 1–649. Available from: https://research.monash.edu/en/publications/cochrane-handbook-for-systematic-reviews-of-interventions-cochran. |
[18] | Study Quality Assessment Tools | NHLBI, NIH [Internet]. [cited 2022 Apr 17]. Available from: https://www.nhlbi.nih.gov/health-topics/study-quality-assessment-tools. |
[19] | Neumann I , Pantoja T , Peñaloza B , Cifuentes L , Rada G . El sistema GRADE: Un cambio en la forma de evaluar la calidad de la evidencia y la fuerza de recomendaciones. Rev Med Chil [Internet]. (2014) [cited 2022 Apr 28]; 142: (5):630–5. Available from: https://pubmed.ncbi.nlm.nih.gov/25427021/. |
[20] | Tufanaru C , Munn Z , Stephenson M , Aromataris E . Fixed or random effects meta-analysis? Common methodological issues in systematic reviews of effectiveness. Int J Evid Based Healthc [Internet]. (2015) [cited 2020 Oct 1];13: (3):196–207. Available from: https://pubmed.ncbi.nlm.nih.gov/26355603/. |
[21] | DerSimonian R , Laird N . Meta-analysis in clinical trials. Control Clin Trials. (1986) ;7: (3):177–88. |
[22] | Higgins JPT , Thompson SG . Quantifying heterogeneity in a meta-analysis. Stat Med [Internet]. (2022) [cited 2020 Nov 7];21: (11):1539–58. Available from: https://pubmed.ncbi.nlm.nih.gov/12111919/. |
[23] | Egger M , Smith GD , Schneider M , Minder C . Bias in meta-analysis detected by a simple, graphical test. Bmj [Internet]. (1997) [cited 2019 Dec 4];315: (7109):629–34. Available from: https://pubmed.ncbi.nlm.nih.gov/9310563/. |
[24] | Lambert JT , Darmahkasih AJ , Horn PS , Rybalsky I , Shellenbarger KC , Tian C , et al. Neurodevelopmental, behavioral, and emotional symptoms in Becker muscular dystrophy. Muscle Nerve. (2020) ;61: (2):156–62. |
[25] | Mori-Yoshimura M , Mizuno Y , Yoshida S , Minami N , Yonemoto N , Takeuchi F , et al. Social involvement issues in patients with Becker muscular dystrophy: A questionnaire survey of subjects from a patient registry. Brain Dev. (2018) ;40: (4):268–77. |
[26] | Mori-Yoshimura M , Mizuno Y , Yoshida S , Ishihara N , Minami N , Morimoto E , et al. Psychiatric and neurodevelopmental aspects of Becker muscular dystrophy. Neuromuscul Disord. (2019) ;29: (12):930–9. |
[27] | Colombo P , Nobile M , Tesei A , Civati F , Gandossini S , Mani E , et al. Assessing mental health in boys with Duchenne muscular dystrophy: Emotional, behavioural and neurodevelopmental profile in an Italian clinical sample. Eur J Paediatr Neurol EJPN Off J Eur Paediatr Neurol Soc. (2017) ;21: (4):639–47. |
[28] | Darmahkasih AJ , Rybalsky I , Tian C , Shellenbarger KC , Horn PS , Lambert JT , et al. Neurodevelopmental, behavioral, and emotional symptoms common in Duchenne muscular dystrophy. Muscle and Nerve [Internet]. (2020) ;61: (4):466–74. Available from: https://pubmed.ncbi.nlm.nih.gov/31909820/. |
[29] | Pane M , Lombardo ME , Alfieri P , D’Amico A , Bianco F , Vasco G , et al. Attention deficit hyperactivity disorder and cognitive function in Duchenne muscular dystrophy: Phenotype-genotype correlation. J Pediatr. (2012) ;161: (4):705–9.e1. |
[30] | Ricotti V , Mandy WPL , Scoto M , Pane M , Deconinck N , Messina S , et al. Neurodevelopmental, emotional, and behavioural problems in Duchenne muscular dystrophy in relation to underlying dystrophin gene mutations. Dev Med Child Neurol. (2016) ;58: (1):77–84. |
[31] | Saito Y , Takeshita E , Komaki H , Nishino I , Sasaki M . Determining neurodevelopmental manifestations in Duchenne muscular dystrophy using a battery of brief tests. J Neurol Sci. (2022) ;440: :120340. |
[32] | Thangarajh M , Hendriksen J , McDermott MP , Martens W , Hart KA , Griggs RC , et al. Relationships between DMD mutations and neurodevelopment in dystrophinopathy. Neurology. (2019) ;93: (17):e1597–604. |
[33] | Fujino H , Saito T , Matsumura T , Shibata S , Iwata Y , Fujimura H , et al. Autism spectrum disorders are prevalent among patients with dystrophinopathies. Neurol Sci Off J Ital Neurol Soc Ital Soc Clin Neurophysiol. (2018) ;39: (7):1279–82. |
[34] | Graus-Porta D , Blaess S , Senften M , Littlewood-Evans A , Damsky C , Huang Z , et al. Beta1-class integrins regulate the development of laminae and folia in the cerebral and cerebellar cortex. Neuron [Internet]. (2001) [cited 2021 Nov 4];31: (3):367–79. Available from: https://pubmed.ncbi.nlm.nih.gov/11516395/. |
[35] | Ash A , Machado L , Raleigh SM , Anthony K . Neuropathophysiology of Duchenne muscular dystrophy: Involvement of the dystrophin isoform Dp71 in cell migration and proliferation. Neuromuscul Disord. (2018) ;28: :S13–4. |
[36] | Reiner O , Karzbrun E , Kshirsagar A , Kaibuchi K . Regulation of neuronal migration, an emerging topic in autism spectrum disorders. J Neurochem [Internet]. (2016) [cited 2021 Nov 4];136: (3):440–56. Available from: https://pubmed.ncbi.nlm.nih.gov/26485324/. |
[37] | Jagadha V , Becker LE . Brain morphology in Duchenne muscular dystrophy: A Golgi study. Pediatr Neurol. (1988) ;4: (2):87–92. |
[38] | Tripp G , Wickens JR . Neurobiology of ADHD. Neuropharmacology [Internet]. (2009) [cited 2021 Nov 4];57: (7– 8):579–89. Available from: https://pubmed.ncbi.nlm.nih.gov/19627998/. |
[39] | Grillner P , Mercuri NB . Intrinsic membrane properties and synapticinputs regulating the firing activity of the dopamine neurons. BehavBrain Res [Internet]. (2002) [cited 2021 Nov 4];130: (1– 2):149–69. Available from: https://pubmed.ncbi.nlm.nih.gov/11864731/. |
[40] | Bruchhage MMK , Bucci M-P , Becker EBE . Cerebellar involvement in autism and ADHD. Handb Clin Neurol. (2018) ;155: :61–72. |
[41] | Vaillend C , Chaussenot R . Relationships linking emotional, motor, cognitive and GABAergic dysfunctions in dystrophin-deficient mdx mice. Hum Mol Genet [Internet]. 2017[cited 2021 Nov 4];26: (6):1041–55. Available from: https://pubmed.ncbi.nlm.nih.gov/28087735/. |
[42] | Domschke K , Lawford B , Young R , Voisey J , Morris CP , Roehrs T , et al. Dysbindin (DTNBP1) - A role in psychotic depression? J Psychiatr Res. (2011) ;45: (5):588–95. |
[43] | Latimer R , Street N , Conway KC , James K , Cunniff C , Oleszek J , et al. Secondary conditions among males with duchenne or becker muscular dystrophy. J Child Neurol. (2017) ;32: (7):663–70. |
[44] | Brown ES , Vera E , Frol AB , Woolston DJ , Johnson B . Effects of chronic prednisone therapy on mood and memory. J Affect Disord. (2007) ;99: (1–3):279–83. |
[45] | Taylor PJ , Betts GA , Maroulis S , Gilissen C , Pedersen RL , Mowat DR , et al. Dystrophin gene mutation location and the risk of cognitive impairment in Duchenne muscular dystrophy. PLoS One. (2010) ;5: (1):e8803. |

