
Influence of polymerized siloxane coating on growth and biofilm formation of aerobic grown nosocomial bacteria
Abstract
A recent study showed that polymerized siloxane (PDMS) coating of biomaterials like polycarbonate, stainless steel or glass results in a hydrophobic and positively charged surface, which is known to be advantageous for cell adhesion. However, when Escherichia coli and Staphylococcus epidermidis were cultured on PDMS coated materials for one hour, this resulted in a significant decrease of bacterial adherence compared to non-coated materials. The study therefore aimed at investigating antimicrobial effects of PDMS. We used nosocomial aerobic grown bacteria S. epidermidis, Pseudomonas aeruginosa, E. coli and the biofilm formation model organism Bacillus subtilis, which were exposed to PDMS in a planktonic culture assay and additionally while biofilm formation. PDMS had a significant impact on the growth of these bacteria in both culture assays. In planktonic culture, PDMS exposition resulted in a decreased growth of all bacteria tested, which was strongly species specific. Biofilm culture in contrast caused an increased growth of E. coli and P. aeruginosa, while growth of S. epidermidis and B. subtilis was decreased. However, these results are based on tests of single species biofilms. Previous to practical application it is necessary to confirm these results by tests in which different bacterial species are able to interact like in natural biofilms resulting in modified bacterial growth behavior and toxin resistance.
1Introduction
Health care-associated infections (HAI’s), or infections acquired in health-care settings are the most frequent adverse event in health-care delivery worldwide. In developed countries 7% of hospitalized patients at any given time, will acquire at least one HAI. In developing countries, case frequency increases onto 10% in hospitalized patients [1]. The main group of HAI’s are those related to the use of invasive devices. 50–70% of the HAI’s reported by the Centers for Disease Control in U.S. are caused by biofilm formation on indwelling medical devices [2, 3]. For this reason, the development of biomaterials and material coating substrates that prevent the formation of biofilms is of great importance to lower the risk of bacterial attachment and growth during and after implantation of a medical device. Recently an aqueous emulsion of polydimethylsiloxane (PDMS), which is a widely used silicone oil, was used as a coating substrate on stainless steel, polycarbonate and soda-lime glass. This coating proved to be non-cytotoxic to eukaryotic cells (fibroblasts), but significantly reduced adherence of aerobic nosocomial bacteria (Escherichia coli and Staphylococcus epidermidis) [4]. However, PDMS provides a hydrophobic surface, which is described to be less resistant to bacterial adhesion than hydrophilic surfaces [5, 6]. For this reason, a study was conducted to clarify whether PDMS can reduce the growth of nosocomial bacteria in a planktonic culture assay. Additionally biofilm growth was studied due to the different behavior of bacteria in planktonic culture and in biofilms [7].
2Materials and methods
2.1Bacteria
The study was performed using Gram-positive bacteria (Staphylococcus epidermidis [ATCC strain-no. 12228], Bacillus subtilis subsp. spizizenii [ATCC strain-no. 6630]), and Gram-negative bacteria (Pseudomonas aeruginosa [ATCC strain-no. 27853; LGC, Germany], Escherichia coli-strain ECM1 which was isolated from C57Bl/6N mouse feces and had proved to not contribute to any diarrheagenic pathovar [4]).
Bacteria were initially cultured on sheep blood agar plates (OXOID, Germany), and after 24 hrs at 36°C transferred to glucose nutrient agar plates (2 vol-%, Sifin diagnostics, Germany), where they were grown for 18 hrs at 36°C before further usage.
2.2Coating substrate
For biofilm assay, we coated polystyrene 96-well microplates with an aqueous emulsion of PDMS (Permanon, Germany). The composition of the PDMS coating substrate is described in more detail elsewhere [1]. For application of the polymerized siloxane, each well was completely filled with an aqueous emulsion of 2.0 vol-% PDMS. After 10 min at 22±2°C the PDMS-solution was exhausted. This coating procedure was repeated three times, resulting in an overall coating time of 30 min.
2.3Biofilm culture assay
This test was performed with single species biofilms using a semi-quantitative biofilm assay as described elsewhere [8, 9]. In brief, one colony of E. coli, one colony of B. subtilis, two colonies of P. aeruginosa, or three colonies of S. epidermidis were inoculated into 50 ml glucose nutrient broth (Sifin diagnostics, Germany) each. Cultures were incubated for 18 hrs at 36°C under continuous shaking (100 rpm). 5μl aliquots of each bacterial culture were transferred into eight wells of a polystyrene-based 96-well microplate. All wells were pre-filled with 200μl DMEM high glucose medium (Sigma Aldrich, Germany). After 18 hrs of biofilm formation at 36°C, bacteria were stained with 0.5% crystal violet (Merck, Germany). Subsequently, cultures were washed three times with H2Odd, followed by destaining with 70% ethanol. Supernatants were transferred to a fresh, uncoated 96-well microplate. Photometrical measurement at 540 nm was performed for quantification of crystal violet.
2.4Planktonic culture assay
5.6 ml of a PDMS in water emulsion in different concentrations (0.2, 2.0, 5.0 and 7.0 vol-%) were added to 74.4 ml glucose nutrient broth to a final volume of 80 ml.
Planktonic growth under the influence of different PDMS concentrations was tested three times separately and in triplicates for each bacterial species. One to three colonies of each bacterium were inoculated into PDMS-supplemented glucose nutrient broth (one colony of E. coli, two colonies of P. aeruginosa, and three colonies of S. epidermidis; depending on individual, species-related different colony sizes). Bacterial growth was quantified using photometric measurement of the optical density (OD) at 590 nm every 30 min for 300 min (550 microplate reader, Bio-Rad, Germany). We excluded B. subtilis from this test due to its strong autoaggregation, which would have resulted in non-assessable measurement inaccuracies.
Because of initial OD reduction in the PDMS-broth mixtures they were pre-incubated for 18 hrs at 36°C under continuous shaking (100 rpm) before inoculation with the bacteria to generate a constant OD.
2.5Statistics
The results were evaluated with the statistics software OriginPro 2016 Version 93G (OriginLab Corporation, Northampton, MA, USA). The significance level was set at 0.05 for all tests performed. The subdivision of the significance levels was defined in * with p≤0.05, ** with p≤0.01 and *** with p≤0.001. Normal distribution was proved by Kolmogorov-Smirnow test. Normally distributed samples could be further investigated with student t-test. Otherwise Welch test was used. For comparison of growth curves, Bonferroni correction was used.
3Results
3.1Effects of PDMS on bacterial growth in planktonic cultures
In planktonic cultures with and without PDMS, the growth curves showed different phases of growth. E. coli almost reached the end of exponential growth or log phase in the examination time period of 300 min. For all bacteria tested, initial growth was retarded due to lag phase, followed by exponential growth. S. epidermidis and especially P. aeruginosa had a slower growth rate than E. coli. After 300 min growth P. aeruginosa was still in the early exponential phase, while S. epidermidis has onward progressed in this phase. None of the bacteria reached the stationary phase. This allowed us to investigate the influence of PDMS on growing cultures and a comparison of effects during growth in a statistical end point analysis.
The impact of PDMS on the growth rate differed between the bacterial species. A dose-dependent effect of PDMS could be shown on E. coli and S. epidermidis as well as for 0.2 to 5 vol-% PDMS on P. aeruginosa. Most commonly, higher concentrations inhibited growth stronger than lower concentrations, except for P. aeruginosa with 7 vol-% PDMS, which reduced bacterial growth less than 5 vol-%.
A 2 vol-% PDMS supplementation to liquid growth medium significantly reduced the growth rate of E. coli, P. aeruginosa and S. epidermidis (p < 0.001; see Fig. 1). Due to the reduced growth of every tested bacterial strain by the used siloxane concentration, we defined 2 vol-% as a sufficient and sensible effective concentration for further tests. After 300 min of incubation in presence of 2 vol-% PDMS the OD of the P. aeruginosa culture was reduced by 89.8% compared to the OD without PDMS (p < 0.001). S. epidermidis showed an OD impairment and growth reduction of 71.5% (p < 0.001) and E. coli exhibited an impairment of 35.0% (p < 0.001). In general, growth of P. aeruginosa was reduced the most, closely followed by S. epidermidis and followed by distance to both by E. coli.
Fig.1
Growth of planktonic bacteria cultures in glucose nutrient broth with 0 vol-% (x), 0.2 vol-% (•), 2 vol-% (♦), 5 vol-% (▴) and 7 vol-% (■) PDMS supplementation; means of three individual tests with triple measurements, each.
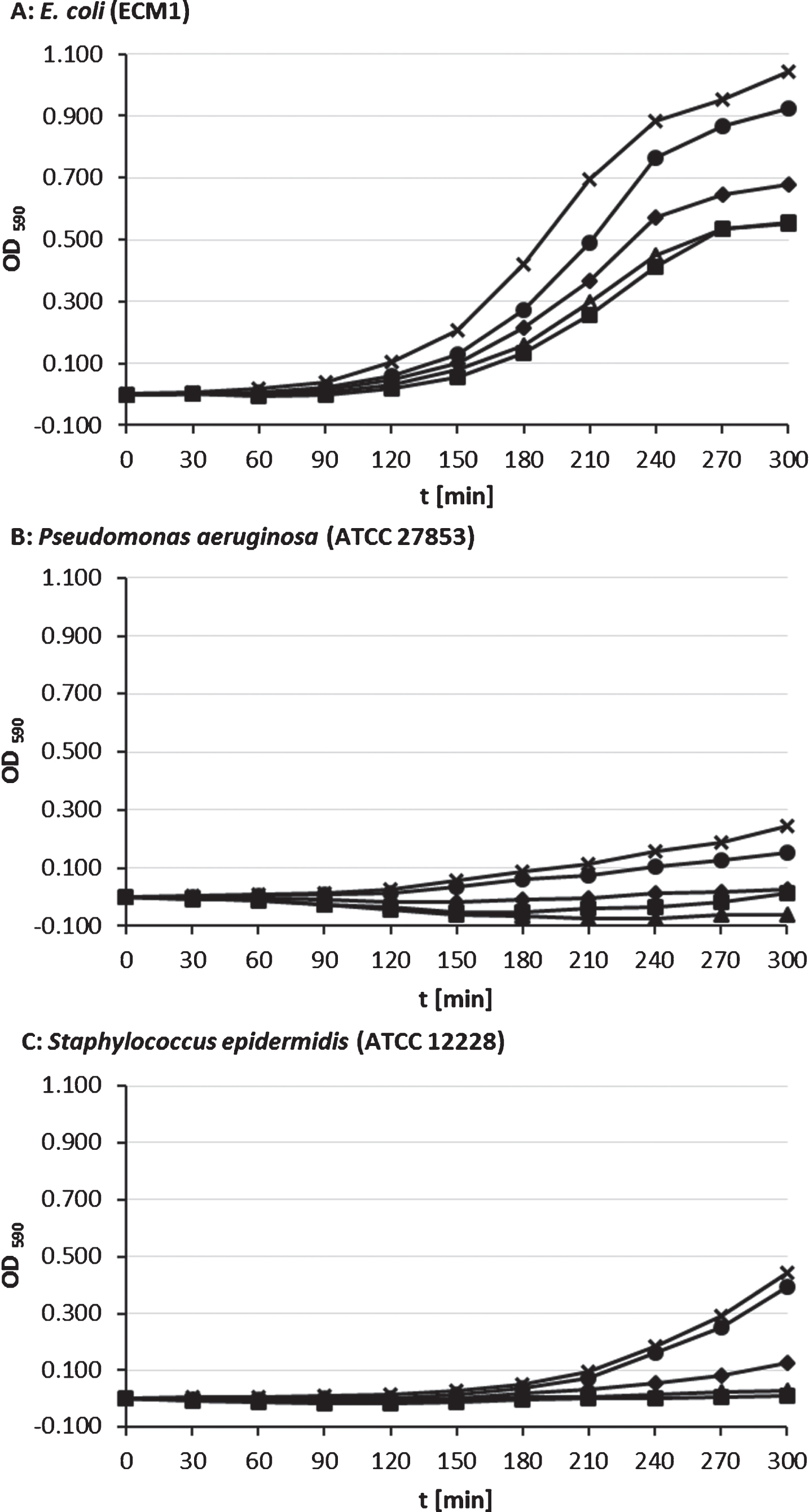
With 7 vol-% supplementation of siloxane maximum growth reduction for S. epidermidis (97.4 %; p < 0.001) and E. coli (45.7%; p < 0.001). For P. aeruginosa we found a maximum growth reduction with 5 vol-% supplemented PDMS (125 %; p < 0.001), whereas bacterial growth with 7 vol-% was less reduced (94.7 %; p < 0.001).
We calculated negative values for optical density for P. aeruginosa and S. epidermidis cultures with 2, 5, and 7 vol-% PDMS, because of blanking against OD of equivalent pre-incubated broth-PDMS mixtures.
3.2Effects of PDMS on bacterial single species biofilm formation and growth
Even without PDMS supplementation cell mass production of the used bacteria differed strongly. While for planktonic cultures the growth rate was the main determining factor, other biofilm-specific factors became important in this assay. Without PDMS exposition S. epidermidis showed the strongest biofilm growth within the time period of testing (18 hrs) followed by P. aeruginosa. Intensity of PDMS-free biofilm growth of E. coli and B. subtilis was similar and much weaker compared to S. epidermidis and P. aeruginosa (see Fig. 2).
Fig.2
Growth of B. subtilis subsp. spizizenii, E. coli, P. aeruginosa and S. epidermidis after 18 hrs of biofilm culture on PDMS-coated/uncoated polystyrene, n = 8.
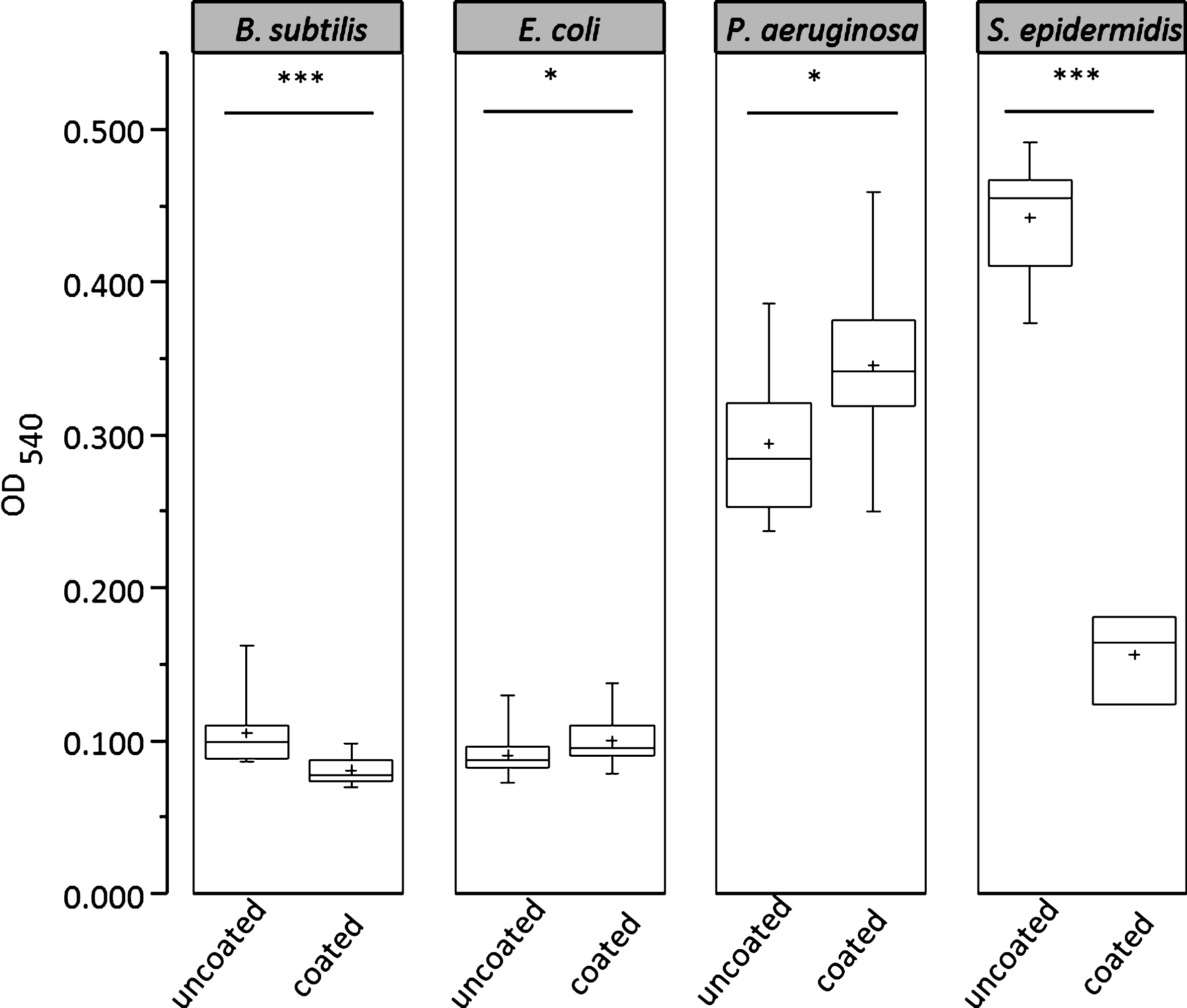
With PDMS exposition, biofilm growth of S. epidermidis and P. aeruginosa was again stronger compared to E. coli and B. subtilis. The impact of PDMS on biofilm production was significant and strongly dependent on the used bacteria. E. coli and P. aeruginosa showed increased biofilm formation on PDMS coated polystyrene, while S. epidermidis and B. subtilis in contrast showed significantly reduced biofilm formation. The bacterial cell mass in biofilms formed by S. epidermidis and B. subtilis was significant reduced by 64.7% (S. epidermidis, p < 0.001) and 23.1% (B. subtilis, p < 0.001) on the PDMS-coated surface, whereas cell mass in biofilms of P. aeruginosa and E. coli was 17.5% (P. aeruginosa, p = 0.011) and 10.1% (E. coli, p = 0.033) higher than without surface coating.
4Discussion
Surface coating for bacterial adherence and growth reduction [3, 10–12] is a promising approach to address the problem of health care-associated infections (HAI’s) which are related to invasive medical devices. A coating based on polymerized siloxanes and establishing a hydrophobic and positively charged surface already proved to reduce the number of adherent aerobic nosocomial bacteria on stainless steel, polycarbonate, and soda-lime glass [4]. The study showed that PDMS also affects the growth rate of aerobic nosocomial bacteria, which were grown planktonically, or in biofilms. This confirms results of a study from Yan and Li [13] which exposed S. epidermidis and P. aeruginosa to silicon oil on blood agar and found PDMS to be antimicrobial. Comparable results were found with anaerobic bacterial strains (Propionibacterium acnes, Peptostreptococcus spp., Bacteroides fragilis, Fusobacterium spp., and Clostridium tertium) [14]. However, in our study different effects of PDMS on bacterial growth in the different culture assays were obvious.
Less intensive growth in tested planktonic bacteria strengthens the growth inhibition due to PDMS. The inhibitory effect of PDMS on bacterial growth seems to be strongly depended on growth rate. P. aeruginosa growth rate is typically slow and we could investigate the strongest inhibitory effect in planktonic cultures onto this bacterial culture. In contrast, growth of E. coli with (a typically) shorter generation time compared to the other bacteria used in this study could not be influenced that intensely.
In planktonic culture with addition of PDMS in different concentrations, even the weakest tested concentration of 0.2% PDMS led to a significant growth reduction of E. coli and P. aeruginosa. Starting from 2 vol-% PDMS growth of S. epidermidis was also significantly inhibited. Effect strength increased with increasing PDMS concentration, implying a dose dependent effect. An exception was P. aeruginosa incubated with 7 vol-% PDMS, because maximum effect for P. aeruginosa was found at 5 vol-% PDMS supplementation.
We calculated negative OD values for some initial points of time in PDMS supplemented cultures of P. aeruginosa and S. epidermidis. This was due to blanking against pre-incubated PDMS-broth mixtures of the same PDMS concentrations. Negative values are based on three additive factors: Slow growing cultures, cytotoxic effects, and putative changing of refraction index due to PDMS and bacterial cell interactions. Temporarily decline of growth curves can be an indicator for loss of bacterial cell integrity. PDMS seems to induce cell death due to cytotoxic effects, delays reproduction and reduces maximum growth rate respectively, as it was shown for silver nanoparticles in E. coli [15]. For P. aeruginosa a cytotoxic effect based on silver that diffused from coating to bacteria was presumed [16].
Biofilms and planktonic cultures of P. aeruginosa differ in gene expression and therefore metabolism like other bacteria, too [17]. Due to optimal growth and reproduction conditions for single cells, metabolism of planktonic log phase cultures is increased compared to biofilms. The physiological state of cells can lead to considerable variation in the receptiveness of bacteria to bactericides [18]. Reduction of viability seems to take place during active metabolic states like growth and cell division. Biofilm-associated tolerance is different from resistance displayed by planktonic cultures of bacteria, because of modified gene expression, which reduces metabolism and growth rate [19]. Therefore, it can be supposed that biofilm specific factors influenced the effect of PDMS-coating on bacterial growth. It is relevant to mention, that we only have examined single-species biofilms. Presumably, combinations of various species like in natural biofilm communities can lead to improved resistance [20].
Traditionally, biofilm formation is divided into at least three steps: Phase of primary attachment, accumulative phase, and disassembly of biofilm structure [21]. Physical forces like van der Waals forces, steric and electrostatic interactions, bacterial appendages like flagella, pili or fimbriae, as well as adhesive surface structures are essential for initial adherence to surfaces and following biofilm formation [22]. S. epidermidis harbors adhesins like the polysaccharide intercellular adhesin (PIA) which is required for enforced biofilm accumulation [21]. Due to hydrophobic and van der Waals interactions it adheres strongly to hydrophobic surfaces like plastics [23]. Autolysin AtlE, a bacterial peptidoglycan-hydrolase, is also of importance for binding on unmodified polystyrene [21]. These characteristics enabled S. epidermidis to be the most potent biofilm producer on plastic surface in our study, and explain why S. epidermidis is one of the most common cause of biofilm-associated opportunistic infections on indwelling medical devices [24]. However, S. epidermidis biofilm growth was most severely affected by 2 vol-% PDMS which led to a reduction of growth for more than 60% compared to the growth without PDMS. In contrast, 2 vol-% PDMS caused an increased biofilm growth of E. coli and P. aeruginosa while P. aeruginosa still exhibiting the stronger growth of both. These results are confirmed by the study of Ozdamar et al. [25] which also approved S. epidermidis less resistant to PDMS than P. aeruginosa. They addressed the effect of 10 vol-% silicone oil on different bacteria and after 12 hrs of growth on brain heart infusion agar at 35°C, S. epidermidis and P. aeruginosa showed an apparent decrease in the number of CFUs, whereby S. epidermidis was able to survive for only 5–7 days and P. aeruginosa for 7–10 days.
In P. aeruginosa at least three polysaccharides (alginate, Pel and PsI) mainly determinate stability of the biofilm structure [22, 26]. Alginate interacts with water and nutrients and supplies nutrients to cells in biofilm [26]. Additionally it reduces the access of toxins like antibiotics to the bacteria and blocks host immune reactions [27].
Strains of E. coli are genotypic and phenotypic heterogeneous [28]. Biofilm formation is mediated through motility by flagella, attachment and microcolony formation by type 1 fimbriae, curli and exopolysaccharides like PGA and colonic acid, antigen 43 as well as other factors for later biofilm maturation [29]. Increased biofilm growth after PDMS exposition might have been caused by the large number of hydrophobic amino acid residues on E. coli flagella, which makes the surface of E. coli very hydrophobic promoting contact of E. coli to hydrophobic surfaces like polystyrene and other polymers [30, 31].
B. subtilis is a robust model organism for examination of biofilm formation of Gram-positive bacteria that manifests in highly structured floating pellicles in liquid cultures [32, 33]. However, laboratory ‘domesticated’ strains of B. subtilis are known to form weaker biofilms in contrast to environmental strains [32]. Also in our study, B. subtilis ssp. spizizenii was a weak biofilm producer. PDMS coating reduced biofilm formation significantly but not as powerful as in S. epidermidis. Additionally it is known that in submerged cultures of B. subtilis BL53 PDMS enhances oxygen supply improving cell metabolism and growth [34]. Therefore the activity of PDMS as oxygen carrier might have been advantageous for the resistance of B. subtilis against the adverse effects of PDMS.
5Conclusion
PDMS had a significant impact on growth of nosocomial bacteria in planktonic and biofilm culture assays. In planktonic cultures, PDMS exposition resulted in a decreased growth of all tested bacteria in a dose-dependent manner in the used 300 min assay. This antimicrobial effect of PDMS was strongly related to the type of bacteria. Biofilm culture resulted in a modified growth behavior and PDMS resistance, whereby the growth of E. coli and P. aeruginosa were not adversely affected any more by PDMS as in the planktonic assay. However, these results are based on tests with single species biofilms. Previous to practical application it is necessary to confirm these results by tests in which different bacteria are able to interact like in natural biofilms, because this can cause modified bacterial growth and resistance [20]. Furthermore, research on the interactions on the molecular level could provide more insight into how antimicrobial effects can be explained and exploited.
Acknowledgments
We would like to thank our technical assistants Ms. Barbara Tüdö and Ms. Undine Ziese and Mr. Rico Eichentopf for technical assistance.
References
[1] | Organization WH. Health care-associated infections fact sheet. (2014) . |
[2] | Bryers JD . Medical biofilms. Biotechnol Bioeng. (2008) ;100: (1):1–18. |
[3] | Darouiche RO . Treatment of infections associated with surgical implants. N Engl J Med. (2004) ;350: (14):1422–9. |
[4] | Pfeiffer S , et al. Influence of polymerized siloxanes on growth of nosocomial aerobic bacteria. Journal of Cellular Biotechnology. (2016) ;2: (1):69–75. |
[5] | Hogt AH , et al. Adhesion of Coagulase-negative Staphylococci to Biomaterials. Microbiology. (1983) ;129: (9):2959–68. |
[6] | Ludwicka A , et al. Attachment of staphylococci to various synthetic polymers. Zentralblatt für Bakteriologie, Mikrobiologie und Hygiene. 1. Abt. Originale. A, Medizinische Mikrobiologie, Infektionskrankheiten und Parasitologie. (1984) ;256: (4):479–89. |
[7] | Penterman J , et al. Rapid evolution of culture-impaired bacteria during adaptation to biofilm growth. Cell Reports. (2014) ;6: (2):293–300. |
[8] | Bangar R , Mamatha B . Biochemical Identification of Enteroaggregative Escherichia Coli among Infants with Acute Diarrhea from Manipal, India. International Journal of Health Sciences. (2007) ;1: (2):237–41. |
[9] | Sarantuya J , et al. Typical enteroaggregative Escherichia coli is the most prevalent pathotype among E.coli strains causing diarrhea in Mongolian children. Journal of Clinical Microbiology. (2004) ;42: (1):133–9. |
[10] | Darouiche RO . Device-associated infections: A macroproblem that starts with microadherence. Clin Infect Dis. (2001) ;33: (9):1567–72. |
[11] | Groll J , et al. Novel surface coatings modulating eukaryotic cell adhesion and preventing implant infection. Int J Artif Organs. (2009) ;32: (9):655–62. |
[12] | Terada A , et al. Bacterial adhesion to and viability on positively charged polymer surfaces. Microbiology. (2006) ;152: (Pt 12):3575–83. |
[13] | Yan H , Li J . An experimental study on antimicrobial activity of silicone oil in vitro. Ophthalmologica. (2008) ;222: (4):245–8. |
[14] | Arici C , et al. An in Vitro Experimental Study on the Antimicrobial Activity of Silicone Oil against Anaerobic Bacteria. Ocular Immunology and Inflammation. (2016) ;24: (2):173–7. |
[15] | Chatterjee T , et al. Antibacterial effect of silver nanoparticles and the modeling of bacterial growth kinetics using a modified Gompertz model. Biochim Biophys Acta. (2015) ;1850: (2):299–306. |
[16] | de Faria AF , et al. Anti-adhesion and antibacterial activity of silver nanoparticles supported on graphene oxide sheets. Colloids Surf B Biointerfaces. (2014) ;113: :115–24. |
[17] | Vital-Lopez FG , Reifman J , Wallqvist A . Biofilm Formation Mechanisms of Pseudomonas aeruginosa Predicted via Genome-Scale Kinetic Models of Bacterial Metabolism. PLoS Comput Biol. (2015) ;11: (10):e1004452. |
[18] | Singh S , et al. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. The Open Microbiology Journal. (2017) ;11: :53–62. |
[19] | Ciofu O , et al. Antibiotic treatment of biofilm infections. Apmis. (2017) ;125: (4):304–19. |
[20] | Wu H , et al. Strategies for combating bacterial biofilm infections. International Journal of Oral Science. (2015) ;7: (1):1–7. |
[21] | Büttner H , Mack D , Rohde H . Structural basis of Staphylococcus epidermidis biofilm formation: Mechanisms and molecular interactions. Frontiers in Cellular and Infection Microbiology. (2015) ;5: :14. |
[22] | Gupta P , et al. Biofilm, pathogenesis and prevention–a journey to break the wall: A review. Arch Microbiol. (2016) ;198: (1):1–15. |
[23] | Shida T , et al. Adherence ability of Staphylococcus epidermidis on prosthetic biomaterials: An in vitro study. Int J Nanomedicine. (2013) ;8: :3955–61. |
[24] | Lassen SB , Lomholt HB , Bruggemann H . Complete Genome Sequence of a Staphylococcus epidermidis Strain with Exceptional Antimicrobial Activity. Genome Announc. (2017) ;5: (10). |
[25] | Ozdamar A , et al. In vitro antimicrobial activity of silicone oil against endophthalmitis-causing agents. Retina (Philadelphia, Pa.). (1999) ;19: (2):122–6. |
[26] | Rasamiravaka T , et al. The Formation of Biofilms by Pseudomonas aeruginosa: A Review of the Natural and Synthetic Compounds Interfering with Control Mechanisms. BioMed Research International. (2015) ;2015: :17. |
[27] | Donlan RM , Costerton JW . Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. (2002) ;15: (2):167–93. |
[28] | Jang J , et al. Environmental Escherichia coli: Ecology and public health implications-a review. J Appl Microbiol. (2017) ;123: (3):570–81. |
[29] | Van Houdt R , Michiels CW . Role of bacterial cell surface structures in Escherichia coli biofilm formation. Research in Microbiology. (2005) ;156: (5):626–33. |
[30] | Vani K , et al. In vitro biocompatiblity of modified polycarbonate as a biomaterial. Colloids Surf B Biointerfaces. (2013) ;108: :191–8. |
[31] | Thevenot P , Hu W , Tang L . Surface chemistry influences implant biocompatibility. Curr Top Med Chem. (2008) ;8: (4):270–80. |
[32] | Mielich-Süss B , Lopez D . Molecular mechanisms involved in Bacillus subtilis biofilm formation. Environmental Microbiology. (2015) ;17: (3):555–65. |
[33] | Vlamakis H , et al. Sticking together: Building a biofilm the Bacillus subtilis way. Nature reviews. Microbiology. (2013) ;11: (3):157–68. |
[34] | de Cesaro A , Da Silva SB , Ayub MAZ . Effects of metabolic pathway precursors and polydimethylsiloxane (PDMS) on poly-(gamma)-glutamic acid production by Bacillus subtilis BL53. Journal of Industrial Microbiology & Biotechnology. (2014) ;41: (9):1375–82. |

