
Inhibition of Tulane Virus replication via exposure to lowbush blueberry (Vaccinium angustifolium) fractional components
Abstract
INTRODUCTION:
Human noroviruses (HuNoVs) are among the leading causes of foodborne illness worldwide and current treatments are ineffective at reducing spread and severity of infection. Previous studies have demonstrated the antimicrobial properties of lowbush blueberries, but little work has been done investigating antiviral properties of blueberries.
OBJECTIVE:
The objective of this study was to determine the antiviral effects of lowbush blueberry fractional components against Tulane Virus in a tissue culture system.
METHODS:
Dried lowbush blueberry powder was separated into five fractional components and LLC-MK2 cells were treated with dilutions of each fraction during virus inoculation to determine if fraction exposure reduced subsequent virus replication.
RESULTS:
Total blueberry extract (F1) exposure resulted in viable particle reduction to undetectable levels (>4.95 log PFU/ml) followed by anthocyanins plus proanthocyanidins (F3; 4.14±0.83 log PFU/ml), monomeric phenolics (F2; 2.90±0.09 log PFU/ml), anthocyanins (F4; 2.27±0.29 log PFU/ml), and proanthocyanidins (F5; 1.73±0.25 log PFU/ml) at a 1 : 2 dilution of each fraction.
CONCLUSION:
The significant reduction of TV caused by blueberry fraction exposure, in combination with no noted signs of cytotoxicity, indicates promising use of blueberry extract antiviral treatment.
1Background
Plants naturally produce a wide variety of metabolites that can serve an important defensive role against bacteria, viruses, and other microbes. Through interactions with pathogenic microorganisms and other biotic factors, such as herbivory, plants have evolved to make a number of phytochemicals that coincidentally have beneficial properties for humans [1, 2]. For example, phenolics derived from Vaccinium species have been experimentally shown to have antibacterial, cardioprotective, neuroprotective, anti-inflammatory, and anti-carcinogenicproperties [3–7]. One species in particular, the lowbush blueberry (V. angustifolium), has distinguished itself from other plants in the genus for its’ berries having high concentrations of antioxidants and phenolics [8]. Previous studies have already demonstrated the inhibitory properties of lowbush blueberry against numerous species of pathogenic bacteria, but there has been comparatively little work looking at their potential antiviral properties [4, 9, 10].
Human noroviruses (HuNoVs) are among the leading causes of foodborne illness worldwide and it is estimated that around 19 to 21 million individuals in the United States alone suffer from norovirus induced gastroenteritis each year [11]. As few as 10–100 viral particles are needed to cause infection and due to having a fecal-oral route mode of transmission, outbreaks generally occur in areas with large numbers of people are in close proximity to each other, such as cruise ships, hospitals, and military bases [12–15]. The most common symptoms of norovirus infection include diarrhea, nausea, vomiting, stomach pain, and dehydration and although generally not lethal, serious complications can occur due to severe dehydration in children, the elderly, and immunocompromised peoples. There is currently no vaccine to prevent human norovirus (HuNoV) infections and the most effective methods to avoid infection are proper sanitation practices.
Being historically uncultivable in a tissue culture or small animal model, the effectiveness of most food-processing and sanitizer treatments at reducing HuNoVs are still relatively poorly understood. Tulane Virus (TV), the prototype strain for the genus Recovirus, is found within the same family as Noroviruses (Caliciviridae) and is becoming an increasingly popular surrogate to use in HuNoV studies [11, 16]. Similarly to HuNoV, the TV genome is organized into three open reading frames, has a fecal-oral route of transmission, and recognize histo-blood group antigens (HGBAs) as receptors [11]. These similarities, in combination with being easily cultivable in a tissue culture system, have arguably made TV currently the most appropriate surrogate to use in HuNoV studies. Other surrogate viruses, such as feline calicivirus (FCV-F9) and murine norovirus (MNV-1) have fallen out of favor of use due to having dissimilar binding receptors and vastly different pathogenesis and infection routes [17].
Given that HuNoV is such a prolific pathogen and treatment is limited to just over-the-counter diarrhea medication, alternative treatment paths need to be investigated [18]. Previous studies have already demonstrated antibacterial and numerous other health properties of lowbush blueberries. However, comparatively little work has been done investigating any potential antiviral activity. Through a highbush blueberry juice treatment, reduction of feline calicivirus (FCV-F9) and bacteriophage MS2 down to undetectable levels after 1 day and 7 days was reported [19]. However, there was only a 1.95 log PFU/ml reduction of MNV-1 after 21 days. Although not of the lowbush species, proanthocyanidins extracted from the leaves of rabbit-eye blueberries (V. virgatum) have demonstrated significant inhibition of hepatitis C RNA expression well below cytotoxic levels [20]. These previous findings suggested that blueberry phytochemicals may have antiviral activity.
2Objective
The objective of this study was to evaluate the ability of blueberry fractional extracts to reduce TV particles in a tissue culture system. Specifically, to determine if TV replication is hindered during simultaneous exposure to lowbush blueberry fractions and host cells.
3Methods
3.1Preparation and quantification of lowbush blueberry fractional components
Following a procedure previously developed to extract blueberry fractions [4], 40 g of freeze-dried lowbush blueberry powder (Durham Research, Londonderry, NH, USA) was separated into five different fractionalcomponents (F1-F5). In short, blueberry powder was stirred overnight in 80% v/v methanol at 25 °C, centrifuged at 5800×g (Eppendorf 5810 R-series, Hamburg, Germany), and then vacuum filtered using No. 3 Whatman paper (Schleicher & Schuell, Maidstone, UK). Excess methanol was then removed by a roto-evaporation (Buchi, Flawii, Switzerland) for 1 h at 45 °C and the remaining solution was placed in a desiccator until completely dry. The extract was suspended in 40 ml of water to make total blueberry extract (F1) and then further separated into monomeric phenolics (F2) and anthocyanins plus pronthocyanidins (F3) using 35-cc Sep-Pak C-18 cartridges (Waters, Milford, MA, USA). LH-20 Sephadex columns (GE Healthcare, Schenectady, NY, USA) were used to separate F3 into anthocyanin (F4) and proanthocyanidin (F5) fractional components using a previously described protocol [21]. After desiccation, each fraction was re-suspended in distilled water and stored at 4°C in the dark until further use.
Three assays used previously [4, 9] to quantify blueberry phenolics were performed. A Folin-Ciocalteu assay [22] was used to quantify total phenolics by comparing samples of each fraction against a standard curve derived from gallic acid solutions. Each sample was read at 725 nm in a 96-well microplate (Biotech, Winoskii, VT, USA) and results were interpreted in equivalents of gallic acid per liter (g/L). Anthocyanin content was measured using a pH differential assay [23] which detects structural differences in the anthocyanin chromophore in solutions of pH 1.0 and 4.5. The difference in absorbencies at 525 and 700 nm read in a 96-well microplate was used as an indication of the relative anthocyanin content and results were reported in cyanidin-3-glucoside equivalents (mg/L). Proanthocyanidin concentration was determined using the 4-dimethylaminocinnamaldehyde (DMAC) protocol [24]. The absorbance read at 640 nm, using a 96-well microplate, of the reaction product between the DMAC reagent (Sigma Aldrich, St. Louis, MO, USA) and the proanthocyanidins were compared to a standard curve derived from procyanidin-A2/DMAC solutions (Infodine, Hillsborough, NJ, USA) and was reported in equivalents of procyanidin-A2 (mM). Methanol was used as the reagent alcohol in the assay instead of ethanol to help stabilize the signal [25]. The pH of each fraction was read at 1 : 1, 1 : 2, 1 : 4, and 1 : 8 dilutions in Media 199 (Corning, Tewksbury, MA, USA) with 10% fetal bovine serum supplementation (FBS; Corning) and 1% penicillin/streptomycin (Corning), here on out referred to as supplemented M199, using an Orion Star A111 pH Meter (Thermofisher, Waltham, MA, USA).
3.2Cultivating Tulane Virus
Tulane Virus (TV), kindly provided by Dr. Jason Jiang at Cincinnati’s Children’s Hospital was cultivated using the LLC-MK2 (ATCC® CCL-7TM) cell line consisting of rhesus monkey (Macaca mulatta) kidney epithelial cells. LLC-MK2 was grown in supplemented M199 at 37 °C in a 5% CO2 atmosphere and after reaching 90% confluency in a tissue culture flask, the old media was aspirated off and the cell monolayer was washed with 10 ml of Dulbecco’s phosphate-buffered saline (DPBS; Corning). Two milliliters of supplemented M199 and 1 mL of TV inoculum (∼5 log PFU/ml) was added to the flask and incubated for 1 h at 37 °C in a 5% CO2 atmosphere. To prevent cell desiccation, the flask was gently rocked throughout the incubation period. After incubation, the media and TV inoculum was aspirated off and replaced with 15 ml of fresh supplemented M199 and the infected cells were then incubated for 48 h at 37 °C in a 5% CO2 atmosphere. Following incubation, the flask was freeze-thawed three times and centrifuged for 3 min at 2500×g and supernatant containing the viral particles were then collected and stored at –80°C until future use. Particle forming units per milliliter (log PFU/ml) was determined by plaque assay and TV harvested were typically in concentrations ∼5 log PFU/ml.
3.3Determination of cytotoxicity of individual blueberry fractions against LLC-MK2 cells
To determine any cytotoxic effects from blueberry fraction treatment, 0.4 ml of blueberry fractions in 1 : 1 dilutions of supplemented M199 were added to individual wells of confluent LLC-MK2 cells in 6-well plates and incubated for 1 h under 5% CO2 at 37 °C. To prevent desiccation the plates were gently rocked during incubation. After 1 h, the fraction solution was then aspirated and treated wells were overlaid with 2.5 ml of an agarose solution consisting of 50% Media 199, 10% FBS, 1% antibiotic/antimycotic solution (Corning), 6% sterile deionized water, and 33% 1.5 w/v agarose (Invitrogen, Carlsbad, CA) and incubated for four days in 5% CO2 at 37 °C. After incubation, cells were fixed with 10% formalin (VWR) and cytotoxicity was determined by visual comparison of the treated cell bed to the control after staining with 0.05% crystal violet (VWR, Radnor, PA) using an inverted compound microscope [26].
3.4Evaluation of growth response of TV to individual blueberry fractions by plaque assay
Serial dilutions of TV particle solutions in supplemented M199 were treated in a 1 : 1 ratio of five fractions in 1 : 1, 1 : 2, 1 : 4, and 1 : 8 dilutions to make final treatment dilutions of 1 : 2, 1 : 4, 1 : 8, and 1 : 16, respectively. Each of the five fractions had four separate concentration treatments consisting of three replicates each. Controls of supplemented M199, without blueberry fraction, and DBPS were included in every trial. Treatment consisted of 0.4 mL of the virus/fraction solution applied to individual wells of confluent LLC-MK2 cells in 6-well plates and incubated for 1 h under 5% CO2 at 37 °C. A 1-h treatment time was selected due to TV being incubated with the LLC-MK2 cells for 1 h during cultivation. After 1 h, the fraction/virus solution was then aspirated and treated wells were overlaid with 2.5 ml of the previously described agarose solution and incubated for four days in 5% CO2 and 37 °C. After incubation cells were fixed with 10% formalin and stained with 0.05% crystal violet. Particle forming units per ml (log PFU/ml) was determined by visually counting plaques in the cell bed.
3.5Statistical analysis
The experiments were repeated three times. TV reduction was determined by comparing the recovered log PFU/ml, as measured by plaque assay, of each dilution of fraction treatment and the media control compared to any reduction seen by the DBPS control. Analysis of Variance (ANOVA) was performed on both the pH and viral reduction data using IBM SPSS Statistics 22 software and significance of differences was defined as P < 0.05. Differences among means were determined using Tukey’s Honest Squared Deviance (HSD).
4Results
4.1Characterization of fractions
Three assays previously used with lowbush blueberries [4, 9] were performed to determine the relative phenolic content of each blueberry fraction and results are recorded in Table 1. The total blueberry extract (F1) had the highest concentration of total phenolics, anthocyanins, and proanthocyanidins while the anthocyanins plus proanthocyanidins (F3) predictably had the second highest concentrations. The monomeric phenolics (F2) showed the lowest concentrations for all three categories of measured phenolics.
Table 1
Analysis of the concentration of lowbush blueberry fractions using folin-ciocalteu, pH differential, and 4-dimethylamiocinnamaldehyde (DMAC) methods. The concentrations are presented as means±standard deviation and consist of three replicates
| Method | Total blueberry extract | Monomeric phenolics | Anthocyanins plus proanthocyanidins | Anthocyanins | Proanthocyanidins |
| Folin-Ciocalteu (gallic acid equivalents (g/L)) | 10.97±5.50 | 2.66±0.60 | 8.79±2.40 | 2.96±1.10 | 3.13±2.13 |
| pH Differential (cyanidin-3-glycoside equivalents (mg/L)) | 99.73±9.90 | 0.43±0.15 | 51.37±0.61 | 48.48±1.93 | 12.53±1.55 |
| DMAC (equivalents of procyanidin-A2 (mM)) | 4.89±0.01 | 0.20±0.01 | 4.76±0.18 | 0.24±0.01 | 0.78±0.01 |
The pH of each fraction was measured in four dilutions of supplemented media 199 and are listed in Table 2. Proanthocyanidins (F5) had the highest pH, outside of the 1 : 8 dilution of F1, and the lowest range of the five fractions at all four dilutions, ranging from 7.43±0.03 to 7.76±0.03. At the lowest dilution, anthocyanins plus proanthocyanidins (F3) had the lowest pH (1.39±0.06) while the anthocyanins (F4) where shown to have the lowest pH at the highest dilution (2.46±0.04). The total blueberry extract had the highest pH range of the blueberry fractions (3.87±0.01 to 7.46±0.01). Except in the cases of the 1 : 1 and 1 : 2 dilutions of F4 and F5, dilution of each fractional components resulted in a significant (P < 0.05) change in pH. Equivalent dilutions of different fraction treatments were all statistically different (P < 0.05) except in the case of the 1 : 1 dilution of F2 and F3.
Table 2
Average pH readings of diluted lowbush blueberry fractional components. Values are presented as means±standard deviation and consist of three replicates. All fractions were diluted in Media 199 (10% FBS, 1% penicillin/streptomycin). Dilutions of same fractions with different letters (e.g. a) or comparable dilutions of different fractions with different letters (e.g. A) are statistically different (P≤0.05)
| Fraction | 1 : 1 Dilution | 1 : 2 Dilution | 1 : 4 Dilution | 1 : 8 Dilution |
| Total blueberry extract | 3.87±0.01aA | 4.40±0.01bA | 6.02±0.00cA | 7.46±0.01dA |
| Monomeric phenolics | 1.47±0.03aBC | 1.67±0.03bB | 2.49±0.01cB | 3.95±0.01dB |
| Anthocyanins plus proanthocyanidins | 1.39±0.06aC | 2.26±0.01bC | 2.87±0.06cC | 3.76±0.06dC |
| Anthocyanins | 1.94±0.02aD | 2.01±0.04aD | 2.23±0.03bD | 2.46±0.04cD |
| Proanthocyanidins | 7.43±0.03aE | 7.48±0.01aE | 7.67±0.02bE | 7.76±0.03cE |
4.2Cytotoxicity of blueberry fractions
LLC-MK2 cells in 6-well plates were treated with dilutions of each blueberry fraction at their lowest dilution for 1 h and then were incubated with an agarose overlay for four days. After the incubation period, the cell beds were compared to a control under an inverted compound microscope for signs of cytotoxicity. There were no signs of cytotoxicity observed for all five fractions, indicating that the treatments were of appropriate concentrations.
4.3Assessment of virus reduction through plaque assay
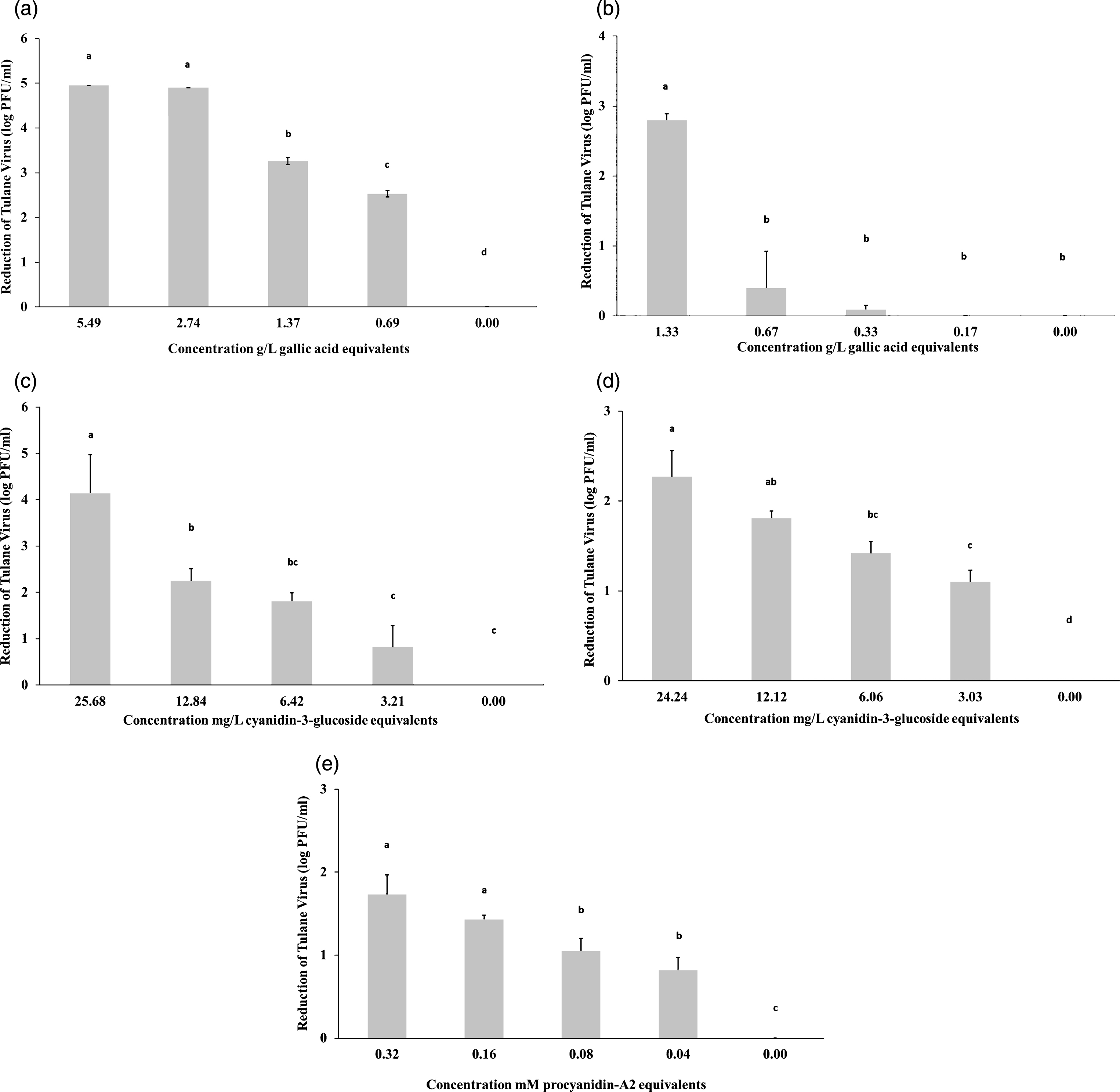
Of the five fraction treatments, F1 treatment resulted in the highest reduction of TV viable particles compared to the control (Fig. 1a). The 1 : 2 and 1 : 4 dilutions, with 5.49 and 2.74 g/L gallic acid equivalents respectively, reduced TV down to undetectable levels (<1 log PFU/ml). At the same dilution of gallic acid equivalents, F2 (Fig. 1b) showed consistently lower levels of reduction compared to F1. The third fraction (Fig. 1c) showed higher reductions than both F4 (Fig. 1d) and F5 (Fig. 1e). Given that both F1 and F3 had higher reductions than their constituents, these indicate that a combination of the various phenolics have a stronger antiviral effect than just one type alone.
Fig.1
The growth response of Tulane virus to total blueberry extract (a), monomeric phenolics (b), anthocyanins plus proanthocyanidins (c), anthocyanins (d), and proanthcyanidins (e). Data are represented as mean±standard deviation and consist of three replicates. Means with different letters are significantly different (P≤0.05) and the detection limit is <1 log PFU/ml.
5Discussion
Although lowbush blueberry has been previously demonstrated to have strong antibacterial properties, to the best of our knowledge, this study is the first time that lowbush blueberry extracts have been directly tested individually for antiviral capabilities. However, prior studies using cranberry juice and extract have beenperformed. Structural changes, albeit minimal, of feline calicivirus particles when treated with cranberry juice and proanthocyanidins has been previously reported [27]. In another study, rotavirus SA-11 examined under TEM after treatment with cranberry juice developed single-shelled virus-like-particles, while the controls had their typical double-shelled formation [28]. Finally, using ELISA and EM imaging, researchers were able to determine that rotavirus lose capsid integrity after treatment with cranberry and grape extracts and suggested that proanthocyanidins (PACs) can potentially block viral bonding to receptors [29].
While we cannot possibly elude any potential mechanism from the present study, there was a clear reduction in new TV particle formation in the presence of the blueberry phenolics. Given that the interaction between host-cell, virus, and fraction was only an hour, it is likely that the blueberry fractions either ruptured the viral particle itself or interfered with the binding to the host cells. Although anthocyanins and proanthocyanidins were selected to be isolated in this study, results from the F-C assay indicate that there are other phenolics present in the fractions that could be contributing to virus inhibition. Additionally, given that the two fractions that had the highest total phenolic content resulted in the strongest reduction and that less inhibition was seen with dilutions of other fractions containing comparable anthocyanin and proanthocyanidin concentrations, it appears that the combination of multiple types of phenolics has a stronger antiviral effect than the individual classes themselves. We hypothesize that different phenolics, may possess different antiviral mechanisms and when combined they possibly may have a more powerful antiviral property.
Regarding pH, previous work [30] reported that TV was stable at a range of pH 3–8, with noticeable reductions in virus titer at pH < 2.5 and >9.0. The highest reduction of TV seen in this study was the total blueberry extract fraction (F1) which was measured to have a pH of 3.87±0.01 (Table 2). The proanthocyanidins (F5) had the overall highest pH with a range of 7.43±0.03 –7.76±0.03. However, when compared with the highest dilution of F1 which had a comparable pH of 7.46±0.01, the reductions seen using the proanthocyanidin fraction were all lower. Fractions 2–4 all had pH values ranging from 1.39–3.76, and yet, only exposure to F3 inhibited TV reproduction more than 3 log PFU/ml. At its lowest dilution, F3 had a pH of 1.39±0.06 which strongly inhibited TV. F2 and F4 had pH’s of 1.47±0.03 and 1.94±0.02 respectively and had comparatively little virus inhibition. Also, statistical differences in pH rarely corresponded to changes in TV reduction. These data indicated that in pH was not a strong factor hindering TV reproduction impediment in this system.
6Conclusions
In conclusion, this study demonstrates that TV particles are sensitive to phenolic produced by V. angustifolium and that exposure to multiple classes of phenolics resulted in higher levels of particle replication inhibition. Thus, blueberry extracts have promise for use as naturally antiviral ingredients and potential food additives. However, further studies must be performed before any claims for blueberry fraction use in humans can be claimed. This study is one contribution to a larger body of research regarding focusing on the benefits of Vaccinium use in human health and although further work in vivo targeted at host-pathogen interactions in the presence of blueberry extracts will be necessary, the outcomes of this study and future work will be valuable in further understanding the health and protective qualities of lowbush blueberries.
Conflict of interest
The authors have no conflict of interest to report.
Acknowledgments
A special thanks goes towards Deborah Bouchard and Dr. Anne Lichtenwalner for assistance with cell culture. Tulane Virus was provided by Cincinnati Children’s hospital. Support for this study was provided by the Wild Blueberry Association of North America and the University of Maine Center for Undergraduate Research.
References
[1] | Neilson EH , Goodger JQ , Woodrow IE , Møller BL . Plant chemical defense: At what cost? Trends Plant Sci (2013) ;18: (5):250–8. |
[2] | Wink M . Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry. (2003) ;64: (1):3–19. |
[3] | Lacombe A , Wu VC , Tyler S , Edwards K . Antimicrobial action of the American cranberry constituents; phenolics, anthocyanins, and organic acids, against Escherichia coli O157: H7. Int J of Food Microbiol. (2010) ;139: (1):102–7. |
[4] | Lacombe A , Wu VC , White J , Tadepalli S , Andre EE . The antimicrobial properties of the lowbush blueberry (Vaccinium angustifolium) fractional components against foodborne pathogens and the conservation of probiotic Lactobacillus rhamnosus. Food Microbiol. (2012) ;30: (1):124–31. |
[5] | Wu VC-H , Qiu X , Bushway A , Harper L . Antibacterial effects of American cranberry (Vaccinium macrocarpon) concentrate on foodborne pathogens. Lwt-Food Sci and Technol. (2008) ;41: (10):1834–41. |
[6] | Zafra-Stone S , Yasmin T , Bagchi M , Chatterjee A , Vinson JA , Bagchi D . Research Article Berry anthocyanins as novel antioxidants in human health and disease prevention. Mol Nutr Food Res. (2007) ;51: :675–83. |
[7] | Côté J , Caillet S , Doyon G , Sylvain J-F , Lacroix M . Bioactive compounds in cranberries and their biological properties. Crit Rev Food Sci Nutr. (2010) ;50: (7):666–79. |
[8] | Wolfe KL , Liu RH . Structure–activity relationships of flavonoids in the cellular antioxidant activity assay. J Agricult Food Chem. (2008) ;56: (18):8404–11. |
[9] | Lacombe A , Tadepalli S , Hwang C-A , Wu VC . Phytochemicals in lowbush wild blueberry inactivate Escherichia coli O157: H7 by damaging its cell membrane. Foodborne Pathog and Dis. (2013) ;10: (11):944–50. |
[10] | Schmidt BM , Howell AB , McEniry B , Knight CT , Seigler D , Erdman JW , Lila MA . Effective separation of potent antiproliferation and antiadhesion components from wild blueberry (Vaccinium angustifolium Ait.) fruits. J Agricult Food Chem. (2004) ;52: (21):6433–42. |
[11] | Li J , Predmore A , Divers E , Lou F . New interventions against human norovirus: Progress, opportunities, and challenges. Annu Rev Food Sci Technol. (2012) ;3: :331–52. |
[12] | Fankhauser RL , Monroe SS , Noel JS , Humphrey CD , Bresee JS , Parashar UD , Ando T , Glass RI . Epidemiologic and molecular trends of “Norwalk-like viruses” associated with outbreaks of gastroenteritis in the United States. Journal Infect Dis. (2002) ;186: (1):1–7. |
[13] | Koopmans M , von Bonsdorff C-H , Vinjé J , de Medici D , Monroe S . Foodborne viruses. FEMS Microbiology Rev. (2002) ;26: (2):187–205. |
[14] | Koopmans M . Progress in understanding norovirus epidemiology. Curr Opin Infec Dis. (2008) ;21: (5):544–52. |
[15] | Teunis PF , Moe CL , Liu P , E Miller S , Lindesmith L , Baric RS , Le Pendu J , Calderon RL . Norwalk virus: How infectious is it? J Med Virol (2008) ;80: (8):1468–76. |
[16] | Drouaz N , Schaeffer J , Farkas T , Le Pendu J , Le Guyader FS . Tulane virus as a potential surrogate to mimic norovirus behavior in oysters. Appl Environ Microbiol. (2015) ;81: (15):5249–56. |
[17] | Li X , Ye M , Neetoo H , Golovan S , Chen H . Pressure inactivation of Tulane virus, a candidate surrogate for human norovirus and its potential application in food industry. Int Journal Food Microbiol. (2013) ;162: (1):37–42. |
[18] | Su XW , Sangster MY , D’Souza DH . In vitro effects of pomegranate juice and pomegranate polyphenols on foodborne viral surrogates. Foodborne Pathog and Dis. (2010) ;7: (12):1473–9. |
[19] | Horm KM , Davidson PM , Harte FM , D’Souza DH . Survival and inactivation of human norovirus surrogates in blueberry juice by high-pressure homogenization. Foodborne Pathog and Dis. (2012) ;9: (11):974–9. |
[20] | Takeshita M , Ishida Y-i , Akamatsu E , Ohmori Y , Sudoh M , Uto H , Tsubouchi H , Kataoka H . Proanthocyanidin from blueberry leaves suppresses expression of subgenomic hepatitis C virus RNA. Journal Biol Chem. (2009) ;284: (32):21165–76. |
[21] | Kalt W , MacKinnon S , McDonald J , Vinqvist M , Craft C , Howell A . Phenolics of Vaccinium berries and other fruit crops. J Sci Foof Agr. (2008) ;88: (1):68–76. |
[22] | Slinkard K , Singleton VL . Total phenol analysis: Automation and comparison with manual methods. Am J Enol Vitic. (1977) ;28: (1):49–55. |
[23] | Lee J , Rennaker C , Wrolstad RE . Correlation of two anthocyanin quantification methods: HPLC and spectrophotometric methods. Food Chem. (2008) ;110: (3):782–6. |
[24] | Payne MJ , Hurst WJ , Stuart DA , Ou B , Fan E , Ji H , et al. Determination of total procyanidins in selected chocolate and confectionery products using DMAC. J AOAC Int. (2010) ;93: (1):89–96. |
[25] | Treutter D . Chemical reaction detection of catechins and proanthocyanidins with 4-dimethylaminocinnamaldehyde. J Chromatogr A. (1989) ;467: :185–93. |
[26] | Su X , D’Souza DH . Grape seed extract for control of human enteric viruses. Appl Enviro Microbiol. (2011) ;77: (12):3982–7. |
[27] | Su X , Howell AB , D’Souza DH . The effect of cranberry juice and cranberry proanthocyanidins on the infectivity of human enteric viral surrogates. Food Microbiol. (2010) ;27: (4):535–40. |
[28] | Lipson SM , Cohen P , Zhou J , Burdowski A , Stotzky G . Cranberry cocktail juice, cranberry concentrates, and proanthocyanidins reduce reovirus infectivity titers in African green monkey kidney epithelial cell cultures. Mol Nutr Food Res. (2007) ;51: (6):752–8. |
[29] | Lipson SM , Ozen FS , Karthikeyan L , Gordon RE . Effect of pH on anti-rotavirus activity by comestible juices and proanthocyanidins in a cell-free assay system. Food Environ Virol. (2012) ;4: (4):168–78. |
[30] | Tian P , Yang D , Quigley C , Chou M , Jiang X . Inactivation of the Tulane virus, a novel surrogate for the human norovirus. Journal of Food Prot. (2013) ;76: (4):712. |


