
Neuroprotective Properties of Eudesmin on a Cellular Model of Amyloid-β Peptide Toxicity
Abstract
Background:
Alzheimer’s disease (AD) is a neurodegenerative disorder characterized by progressive cognitive impairment and memory loss. One of the hallmarks in AD is amyloid-β peptide (Aβ) accumulation, where the soluble oligomers of Aβ (AβOs) are the most toxic species, deteriorating the synaptic function, membrane integrity, and neuronal structures, which ultimately lead to apoptosis. Currently, there are no drugs to arrest AD progression, and current scientific efforts are focused on searching for novel leads to control this disease. Lignans are compounds extracted from conifers and have several medicinal properties. Eudesmin (Eu) is an extractable lignan from the wood of Araucaria araucana, a native tree from Chile. This metabolite has shown a range of biological properties, including the ability to control inflammation and antibacterial effects.
Objective:
In this study, the neuroprotective abilities of Eu on synaptic failure induced by AβOs were analyzed.
Methods:
Using neuronal models, PC12 cells, and in silico simulations we evaluated the neuroprotective effect of Eu (30 nM) against the toxicity induced by AβOs.
Results:
In primary cultures from mouse hippocampus, Eu preserved the synaptic structure against AβOs toxicity, maintaining stable levels of the presynaptic protein SV2 at the same concentration. Eu also averted synapsis failure from the AβOs toxicity by sustaining the frequencies of cytosolic Ca2+ transients. Finally, we found that Eu (30 nM) interacts with the Aβ aggregation process inducing a decrease in AβOs toxicity, suggesting an alternative mechanism to explain the neuroprotective activity of Eu.
Conclusion:
We believe that Eu represents a novel lead that reduces the Aβ toxicity, opening new research venues for lignans as neuroprotective agents.
INTRODUCTION
Alzheimer’s disease (AD) is the most prevalent cause of dementia and neurodegenerative disorders, which are clinically characterized by memory loss and cognitive impairment [1–3]. The histopathological hallmarks of AD are related to protein accumulation of intracellular hyperphosphorylated tau protein and extracellular amyloid-β peptide (Aβ) deposits [4, 5]. The aggregation of Aβ peptide into senile plaques has been considered a major route for the development of AD. The Aβ peptide is produced by the cleavage of amyloid-β protein precursor (AβPP) by β-secretase and γ-secretase [6]. These cleavage products are imprecise, resulting in different Aβ species that can self-aggregate leading to several structures such as protofibrils, fibrils, or plaques. The aggregation process consists of an initial nucleation phase where the monomers undergo a structural change from alpha-helix to beta-sheet, which induces aggregation. This second stage is the elongation phase, where trimers, oligomers, protofibrils, fibrils, and finally, plaques of fibrillar aggregates or senile plaques are formed [7–9]. The γ-secretase product at position 40 (Aβ1-40) and 42 (Aβ1-42) has hydrophobic and fibrillogenic properties and are the most abundant species [9]. The senile plaques are mainly composed of Aβ1-40 [10, 11], which are very toxic species and able to form pores in the cellular membrane, allowing a disruption in ionic homeostasis [12, 13]. The membrane perforation induced by Aβ is associated with an increase in intracellular calcium levels [14], and as demonstrated more recently, with increased passage of ions and molecules as large as glucose and ATP through cell membranes [8, 11, 15, 16]. The increase in extracellular ATP can lead to the activation of P2X purinergic receptors in neurons and glia [17]. These receptors are permeable to Ca2+ and can induce the activation of microglia and mitochondrial dysfunction. These events are known to induce synaptic failure and neuronal death, facilitating neurodegeneration [15, 18–20]. During this process, several neurodegenerative events occur, including neuronal dysfunction, microglial response, astrogliosis, and apoptosis [18, 21, 22].
Several hypotheses have been proposed to understand the origin of AD, including malfunction of the cholinergic system [23], extracellular depositions of Aβ peptide [24], hyperphosphorylation of tau protein in the intracellular space [25], and an increase in oxidative stress [26]. However, most treatments to control AD are focused on cholinergic therapies (i.e., acetylcholinesterase inhibitors) [27]. Galantamine is an alkaloid obtained from Galanthus nivalis showing activity as an acetylcholinesterase inhibitor and nicotinic positive allosteric modulator. This alkaloid has been used to treat cognitive impairment since the cholinergic theory for AD was established [28, 29]. Moreover, other plant metabolites have also been reported as attractive neuroactive compounds with anti-inflammatory, analgesic, or antioxidant properties [8, 30].
Natural products, including polyphenols and alkaloids, have several effects that can potentially benefit central nervous system (CNS) function. For instance, some natural products exhibit antioxidant properties and anti-inflammatory activity, contributing to their neuroprotective effects [17, 30, 31]. Benzofurans are biomedically relevant scaffolds and have been reported as inhibitors of the Aβ peptide fibrillation process, suggesting these chemical cores could help control neurodegenerative disorders such as AD [31–35]. Recently, our team has demonstrated the biological and neuroprotective activity of fomannoxin, the most prevalent natural benzofuran isolated from the Russulale fungus, Aleurodiscus vitellinus, an edible mushroom native to Andean-Patagonian forests. This benzofuran can preserve synaptic structure and function, illustrated by experiments where 10–6M of fomannoxin can induce a reversion or protection against Aβ soluble oligomers and disrupts their aggregation [36]. Lignans isolated from conifers species have been recently studied and shown to exhibit similar anti-inflammatory, antihypertensive, and antioxidant properties [37–40]. Additionally, these privileged natural scaffolds have been studied as potential phytohormones for, among others, the treatment or prevention of cancer and neuroprotective properties [38]. Different type of lignans have been isolated from almost all vascular plants, but their specific role is not entirely understood [41]. It has been suggested that they are part of the plants’ defense system against pathogens [2, 3].
Eudesmin (Eu) (Fig. 1A) is a tetrahydrofurofuranoid lignan that has been isolated from numerous medicinal plants and shown several biological activities including cytotoxic, antibacterial, antifungal, anti-inflammatory, immunostimulant, and inhibitory effects on tumor necrosis factor-alpha (TNF-α) production as well as improving the neurite outgrowth [11, 39, 40, 42, 43]. Moreover, Eu has been reported mainly from the genus Artemisia, exhibiting nitric oxide (NO) inhibitory effects and cytotoxic activities [39, 40]. In the present study, we use Eu isolated from the wood of Araucaria araucana, a Chilean native species to evaluate its neuroprotective abilities using an in vitro model of AD with Aβ soluble oligomers treatments [41]. We investigated the cells viability response, presynaptic preservation, and established a direct relationship with calcium transient recordings and presynaptic markers. This experimental evidence suggests an alternative mechanism to explain the neuroprotective activity of Eu, evaluating the effect of this lignan on the interaction with Aβ during the aggregation process considering in vitro assays and in silico models. Computational studies then indicated the specific residues involved in the interaction between Eu and Aβ.
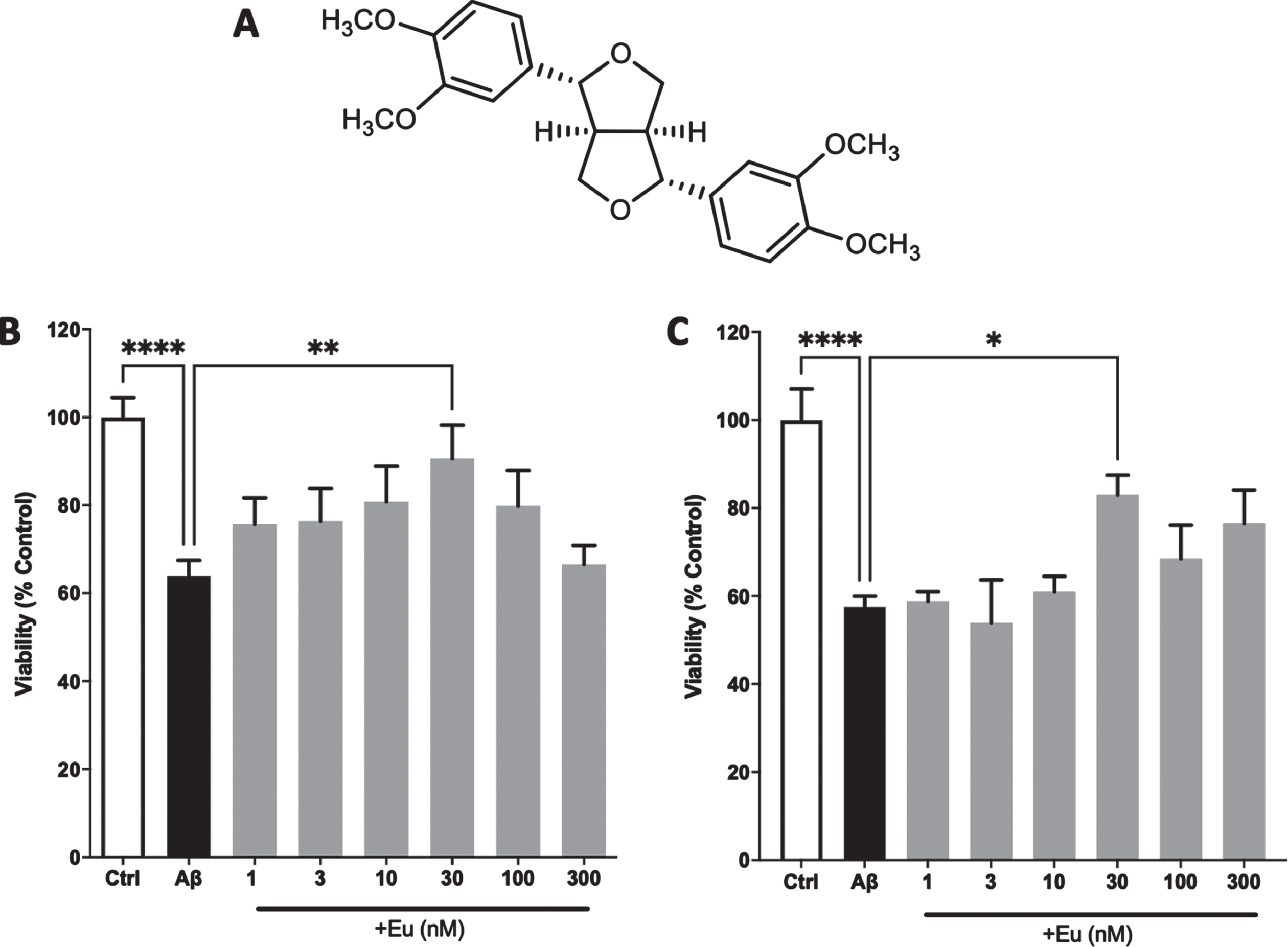
Fig. 1
Neuroprotective effect of Eudesmin against AβOs. Chemical structure of Eudesmin (A). PC12 cells (B) and cortical neurons (C) were co-incubated for 24 h with AβOs (0.5μM) and increasing concentrations of Eudesmin (Eu 1–300 nM). Cellular viability was measured by MTT assay. Values are expressed as the percentage of the control group (without AβOs treatment). *p < 0.05, **p < 0.01, ****p < 0.0001. n = 3.
MATERIALS AND METHODS
PC12 cells culture
PC12 cells were cultured in DMEM 1X (Corning, USA) with 4.5 g/L glucose, L-glutamine, and sodium pyruvate supplemented with 5% fetal bovine serum (Gibco, USA), 5% horse serum (HyClone, USA), and 1% penicillin/streptomycin (Gibco, USA). The cells (passage 20) were incubated under standard conditions (37°C, 5% CO2), plated at a concentration of 50,000 cells/well for experiments and used 24 h after.
Primary cultures of mouse hippocampal neurons
Hippocampal neurons were obtained from 18-day pregnant mouse embryos C57BL/6J following ethical regulations established by NIH and University of Concepción. The cells were plated at density of 320,000 cells/ml. The cells were cultured in DMEM supplemented with 10% horse serum (HyClone, USA), 4μg/ml DNAase (HyClone, USA), and L-glutamine (HyClone, USA). Plating media was replaced after 24 h by DMEM supplemented by 5% horse serum (HyClone, USA), 5% fetal bovine serum (Gibco, USA), and 0.5 % N3 (a mixture of defined nutrients). Hippocampal neurons were maintained under standard conditions (37°C, 5% CO2) in a thermo-regulated incubator. All experiments were performed at 9-10 days in vitro (DIV).
Eudesmin obtention and isolation
Eu extract were obtained from knots of Araucaria araucana (Molina) K. Koch collected in May 2018 on Nahuelbuta range (Biobío Region, 37°41’′43.56”S73°07’49.34”W). Samples were identified by the botanist Dr. Roberto Rodríguez from the University of Concepcion. 100 g of A. araucana wood were chopped, dried at 40°C for 8 h, milled to 1–3 mm. Soxhlet extraction was made with 2–5 g of sample placed in a cellulose thimble with acetone (250 mL) and refluxed for 12 h. The organic layer was filtered through a GV Durapore filter (0.22μm pore size, 13 mm diameter, Millipore, Bedford, MA, USA) and evaporated under reduced pressure, and dried in vacuum to give the extract. Subsequently, isolation of Eu (yield 1% w/w) was obtained after dissolve the extract in hot ethanol (70°C) and kept in cold in 4°C for 8 h. Eu was frozen at –20°C until use with a 96–98% purity as previously described [41].
Preparation of Eudesmin stock
10 mg of Eu extract were dissolved in 2587.69μL DMSO and stored in aliquots at 10 mM. The aliquots where then diluted in DPBS 1x (Gibco, USA) leaving a stock of 1 mM. Four serial dilutions were made in DPBS 1X (Gibco, USA) to achieve the experimental conditions. All the experiments with Eu were at a concentration of 1, 3, 10, 30, 100, 300, and 1000 nM.
Peptide Aβ1-40 preparation
Human synthetic Aβ1-40 peptide (GenicBio, Shanghai, China) was reconstituted in DMSO to a concentration of 2.3 mM and stored at –20°C. Subsequently, the peptide was aggregated in sterile 1X Phosphate-Buffered Saline (Corning, USA) at 80 mM. For the formation of oligomers, Aβ1-40 was stirred vertically at 500 rpm to 37°C for 4 h in a Thermomixer Compact (Eppendorf AG, Hamburg, Germany). All treatments with Aβ1-40 oligomers (AβOs) were made at a final concentration of 0.5 mM.
Cellular viability
Cellular viability changes were evaluated by the in vitro MTT assay kit (Sigma-Aldrich, USA) that measures the ability of mitochondria to reduce 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT salt) to formazan, which has a purple color. The cells were subjected to 1, 3, 10, 30, 100, 300, and 1000 nM of Eu, and then incubated for 2 h in MTT (1 mg/ml). The insoluble formazan was solubilized in 100μL of isopropanol, and the absorbance was read in a NOVOstar multiplate reader at 570 nm (MG Labtech, Germany).
Immunofluorescence
Control and treated hippocampal neurons were fixed with 4% paraformaldehyde for 15 min at 4°C. Afterward, the cells were permeabilized and blocked with 10 % horse serum (HyClone, USA) plus 0.1 % Triton X-100 for 15 min at room temperature. The neurons were subsequently incubated with monoclonal anti-SV2 antibody (1:200, Hybridoma Bank) and polyclonal anti MAP2 (1:200, Synaptic Systems) for 1 h at room temperature (RT) followed by incubation with the corresponding second antibody conjugated to AlexaFluor 488 and Cy3 (Jackson ImmunoResearch, USA) for 1 h at RT. Finally, the samples were mounted in Fluorescent Mounting Media (Dako, Agilent, USA) and the images were obtained using a laser scanning microscope LSM780 (63X oil immersion objective, Zeiss, Germany). Image processing was made with Image J (NIH, Bethesda, MD, USA).
Western blot
Equal amounts of protein obtained from hippocampal neurons treated with AβOs (0.5μM), Eu (30 nM), and AβOs plus Eu (AβOs+ Eu) for 24 h, were separated on 10% SDS-polyacrylamide gels. Protein bands were transferred into PVDF membranes (250 mA, 120 min), blocked with 5% nonfat milk, and incubated with primary antibodies using the following concentrations: 1:1000 anti-SV2 (mouse; Hybridoma Bank; USA) and 1:1000 β-actin (mouse; Santa Cruz Biotechnology, USA). A secondary HRP anti-mouse antibody was used at 1:5000 (Santa Cruz Biotechnology, USA). Immunoreactive bands were revealed using Clarity TM Western ECL substrate (Bio-Rad Laboratories, USA) and visualized with an Odyssey FC detection system (Li-Cor, Lincoln, NE, USA). Quantification of the signals was made with the Image Studio program (Image Studio Inc, USA) and β-actin were used as an internal control for the experiments and quantification.
Calcium imaging
Neurons were treated with AβOs (0.5μM), Eu (30 nM), and AβOs + Eu for 24 h and were loaded with 1μM Fluo-4AM; (Invitrogen, Carlsbad, CA, USA) for 20 min al 37°C, washed 2 times with normal external solution and the mounted on an inverted fluorescent microscope Nikon TE-2000-U (Nikon, Japan). The cells were briefly illuminated (200 ms) and the regions of interest (ROI) were selected on neuronal soma emitting Fluo-4 fluorescence (ex 480 nm, em 510 nm). Images were collected at 1 s intervals during a continuous 200-s exposure using an EMCCD iXon- 16-bit camera (Andor, Northern Ireland). The data were analyzed with the GraphPad Prism program (GraphPad, San Diego, CA, USA).
Thioflavin T fluorescence assay
Aβ1-40 aggregation assay was performed on a 96-microwell plates in the presence of Eu (30 and 300 nM) with 20μM Thioflavin T (ThT, ex: 440 nm, em: 485 nm, Sigma-Aldrich). The ThT fluorescence intensity of the aggregation process of Aβ was measured every 3 min for 4 h using a NOVOstar multiplate reader (MG Labtech, Germany). The plate was kept at 37°C with an orbital agitation of 500 rpm.
In silico prediction (molecular docking simulations)
Protein-ligand docking was performed using the structure of the Aβ1-40 monomer from the Protein DataBank (PDB ID: 1AML) obtained by NMR. The Aβ1-40 structure was prepared to incorporate hydrogens, assign bond orders, fill in missing side chains, and generate protonation states at pH 7±0.2 using Maestro (Schrödinger, LLC, New York, NY, USA). The structure of Eu was obtained in the PubChem database (CID: 234823) and prepared using LigPrep (Schrödinger, LLC) before docking protein-ligand [44]. Initially, a docking protein-ligand was performed using Autodock Vina [45] in which the monomer Aβ1-40 was used as the protein target. Based on these preliminary results, a site-directed docking was performed with Glide (Schrödinger, LLC, New York, NY, 2018) using a receptor grid of 20Å centered on the N-terminal amino acids, with an extra precision (XP) configuration. Analysis of the interface Aβ-Eu included structural and energetic parameters performed by the same software. Additionally, a theoretical MM-GBSA ΔG bind was calculated using Prime (Schrödinger, LLC). All images were created using PyMol (version 1.5, DeLano Scientific LLC).
Data analysis
Statistical analyses were performed using GraphPad Prism (GraphPad, San Diego, CA, USA). The values are expressed as mean±S.E.M. for 3 or more independent experiments and as a percentage of the control (without treatment). Statistical differences were determined using T Student’s test or One-way ANOVA with Tukey or Kruskal Wallis post-test. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001 was considered statistically significant.
RESULTS
Eudesmin has neuroprotective effects against Aβ oligomers on neuronal models
To determinate the neuroprotective effects Eu against AβOs, PC12 cells and hippocampal primary neurons were co-incubated with AβOs, during 24 h (0.5μM) in the presence of Eu (1, 3, 10, 30, 100, and 300 nM) (Fig. 1A). The cellular viability was measured using MTT assay. Figure 1B and 1C show that AβOs significantly reduced cellular viability in PC12 cells (36%) and cortical neurons (42%), respectively. Eu (30 nM) exerted a significant neuroprotective effect on PC12 cells, increasing the viability by 25.4% over control experiments (Fig. 1B). Equally, cortical neurons co-treated with Eu (30 nM) underwent an increase in their viability by 26.7% (Fig. 1C). Furthermore, we also observed that Eu concentrations have no intrinsic toxic properties on our cellular and neuronal models (Supplementary Figure 1).
Eu preserve the synaptic structure by conservation of exocytotic SV2 protein
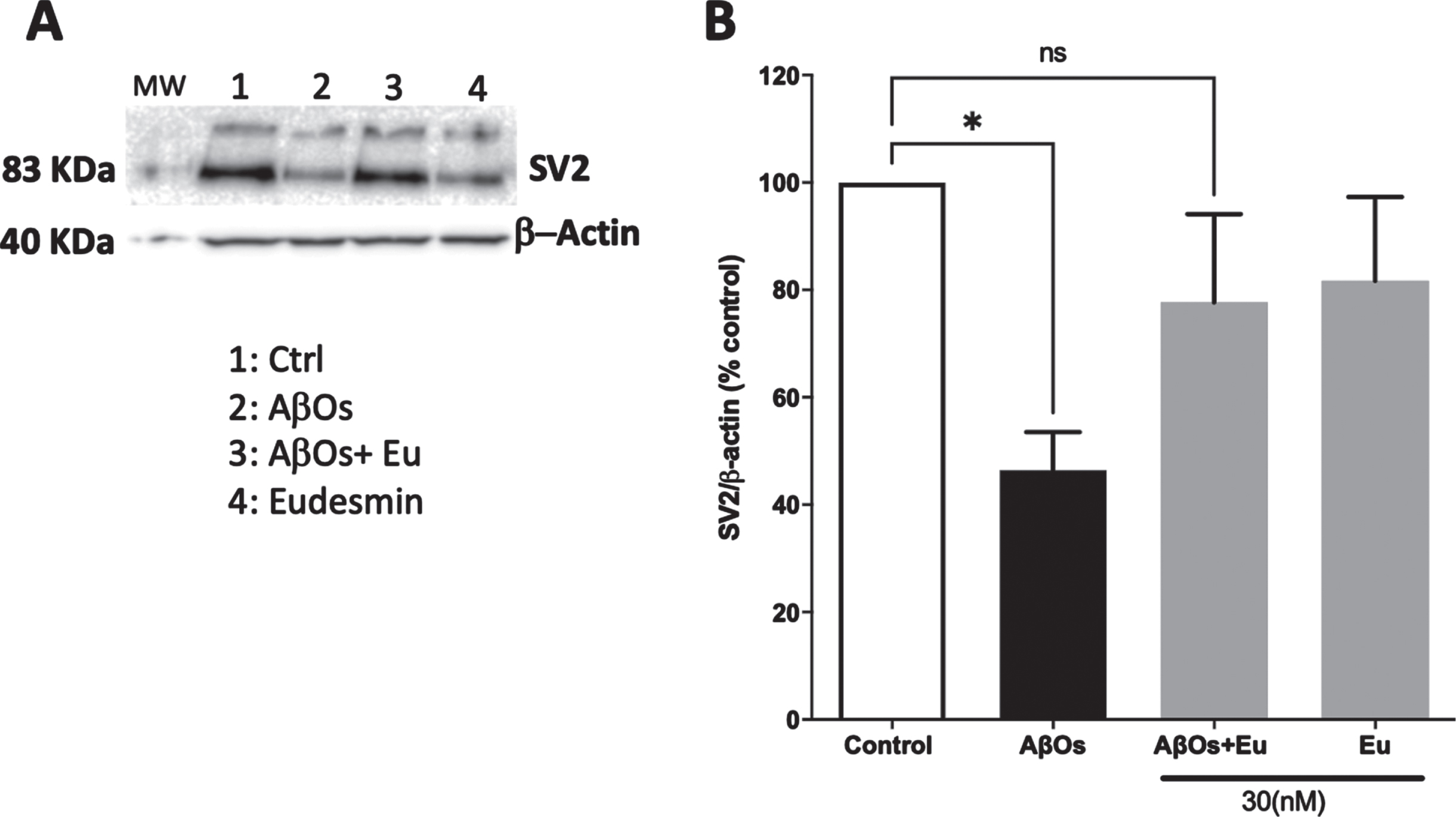
In order to establish a correlation between neuronal viability and synaptic function, we initially analyzed the SV2 immunoreactivity using Eu (30 nM) and AβOs treatments (Fig. 2A) by immunofluorescence experimental approaches, using confocal microscopy techniques. The total fluorescence intensity of SV2 was quantified. The SV2 immunoreactivity showed a significant decrease of 34.5%, in AβOs-treated neurons (Aβ: 65.5±7.30% of the control) (Fig. 2B). These results correlate with our observations in viability studies and suggest that part of this neuroprotective effects may be associated to synaptic function preservation. Eu treatments showed fluorescence intensity similar to controls (Fig. 2B). Moreover, western blot analysis of SV2 signal (Fig. 3A) rendered similar results. AβOs treatments showed a significant decline in SV2 band intensity, 53.5% (Aβ: 46.5±7.05% of the control). Eu co-treatment prevents this decrease (Fig. 3B).
Fig. 2
Neuroprotective effect of Eudesmin on synaptic markers. Immunofluorescence confocal images of hippocampal neurons co-incubated for 24 h with AβOs (0.5μM) and Eudesmin (Eu 30 nM) to analyze MAP2 (red) and SV2 (green), a zoom of each treatment was represented (A). Quantification of total fluorescence SV2 intensity was measured using Image J software (B). Values are expressed as the percentage of the control group (without AβOs treatment). Values are mean±SEM, n = 3 using one-way ANOVA and Bonferroni test. *p < 0.05.
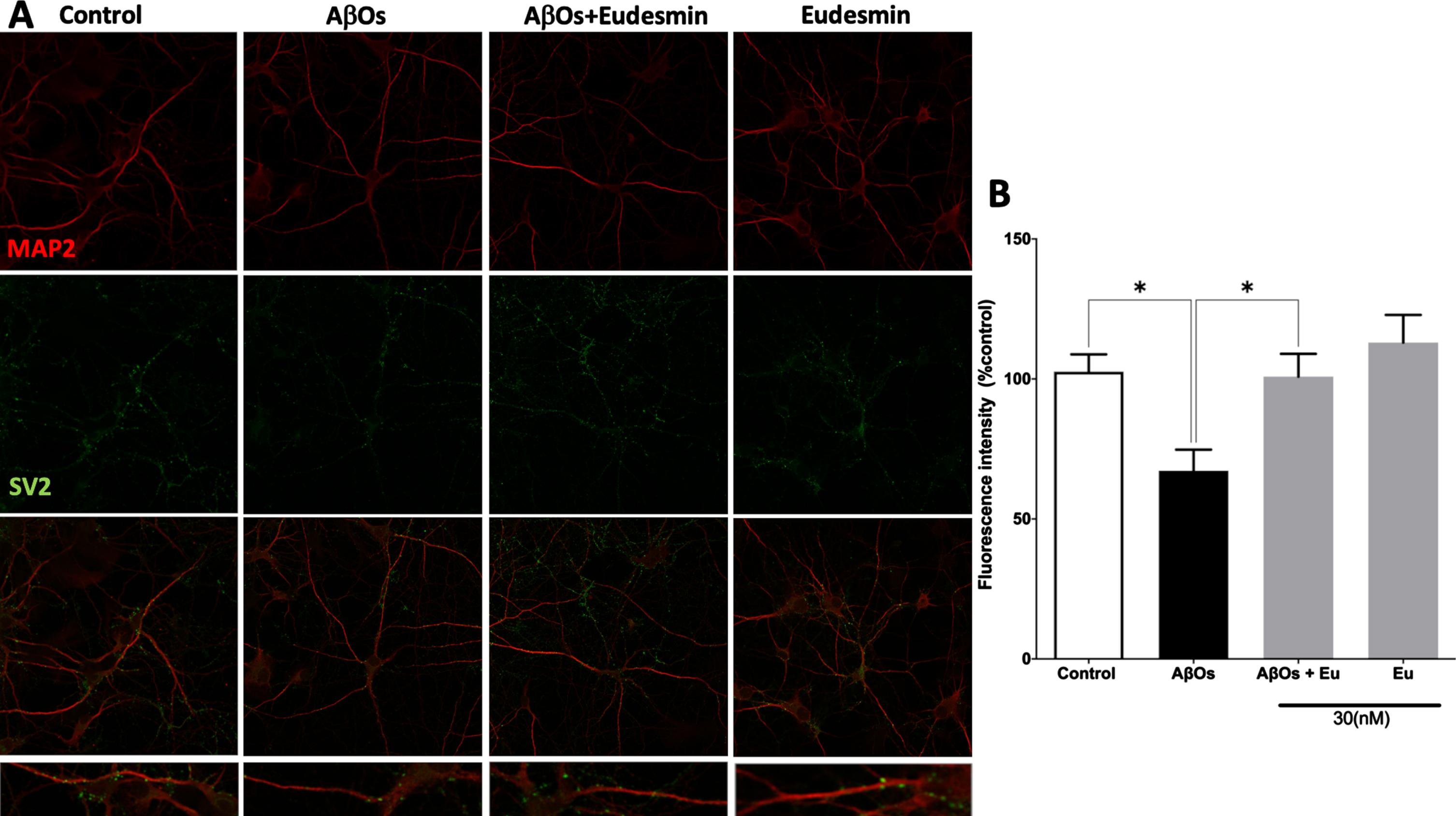
Fig. 3
Neuroprotective effect of Eudesmin on synaptic markers: SV2 detection. Western blot images of neuronal cultures co-incubated for 24 h with AβOs (0.5μM) and Eudesmin (Eu 30 nM) SV2 total expression (A). Total expression intensity was measured using the Odyssey FC detection system. β-actin detection was used as reference. Values are expressed as the percentage of the control group (without AβOs treatment) (B). Values are mean±SEM, n = 3 using one-way ANOVA and Bonferroni test. *p < 0.05 versus control group.
Eudesmin’s synaptic preservation against AβOs is related to Ca2+ signals conservation
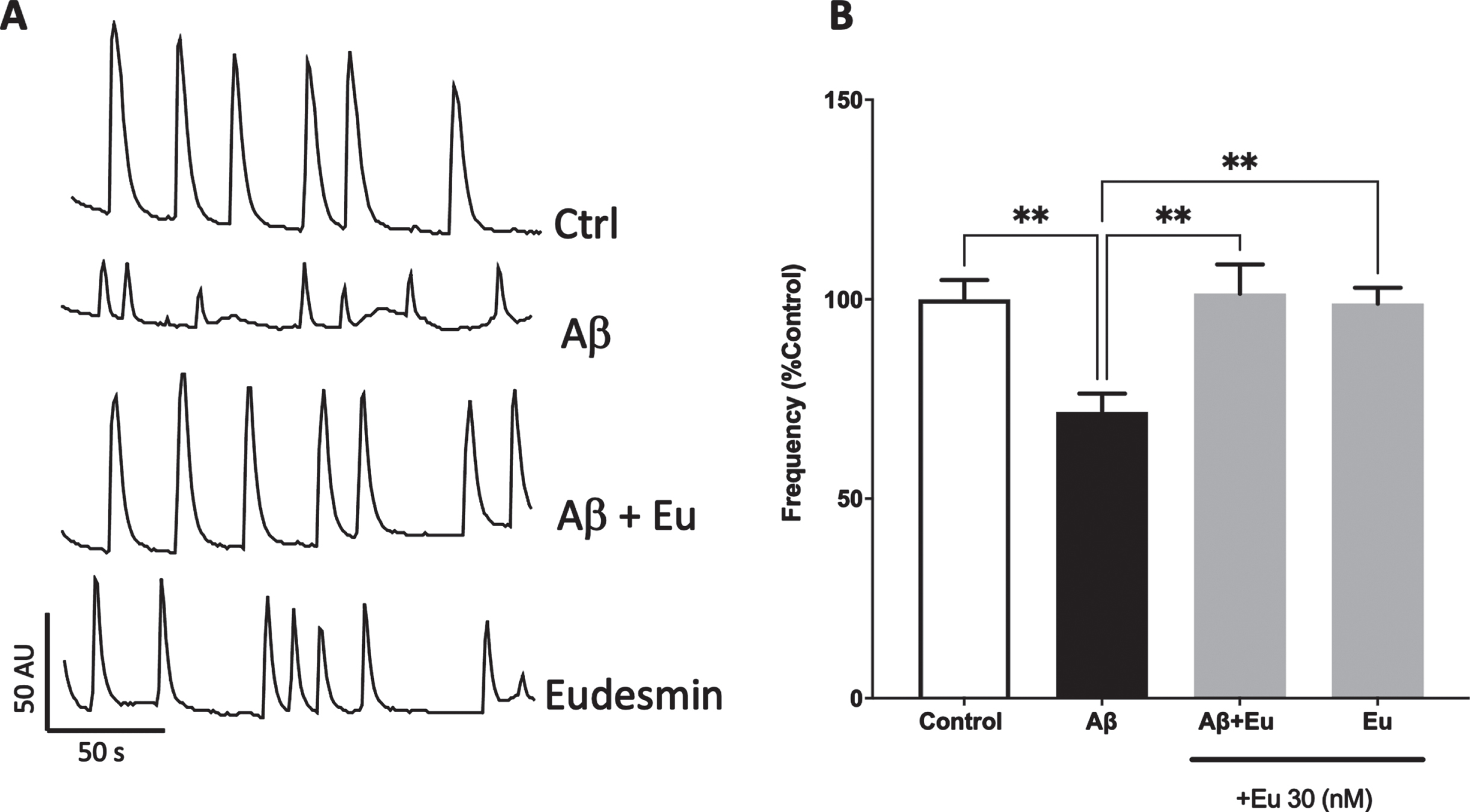
To reinforce our previous results and establish a deeper understanding of the observed neuroprotective properties of Eu, we decided to study the synaptic function preservation by means of intracellular spontaneous Ca+2 signals, using hippocampal neurons in the presence of AβOs and co-treated with Eu (30 nM) (Fig. 4A). After 24 h, we evaluated calcium transients and quantified the frequency of Ca+2 spikes for 200 s (Fig. 4B). AβOs treatments reduced the spikes frequency approximately 30% (Aβ: 71.8±4.54% of the control), in accordance with our previous work [19, 46, 47]. Co-incubation with Eu leads to the preservation of calcium transient’s frequencies on neurons. This result suggests that the synaptic structural network conservation is associated with the preservation of functional synapses. Moreover, Eu has no toxic effects on synaptic communication after treatments and was able to recover the reduction induced by AβOs respect to control condition (Aβ+Eu: 101±7.28%; Eu: 98.9±3.97%) (Fig. 4B).
Fig. 4
Eudesmin restored calcium transients in neurons treated with AβOs. Hippocampal neurons were co-incubated for 24 h with AβOs (0.5μM) and Eudesmin (30 nM). Calcium transients were measured using Fluo-4AM for 200 s. Representative images of calcium transients in cells treated to the different indicated treatments (A). Quantification of calcium transient’s frequency. Percentage values are expressed as the percentage of the control group (without AβOs treatment) (B). Values are mean±SEM, n = 3 using Kruskall-Wallis test, with Dunn’s post-test correction. **p < 0.01.
Eudesmin induces alterations on AβOs aggregation pattern and reduce their toxicity
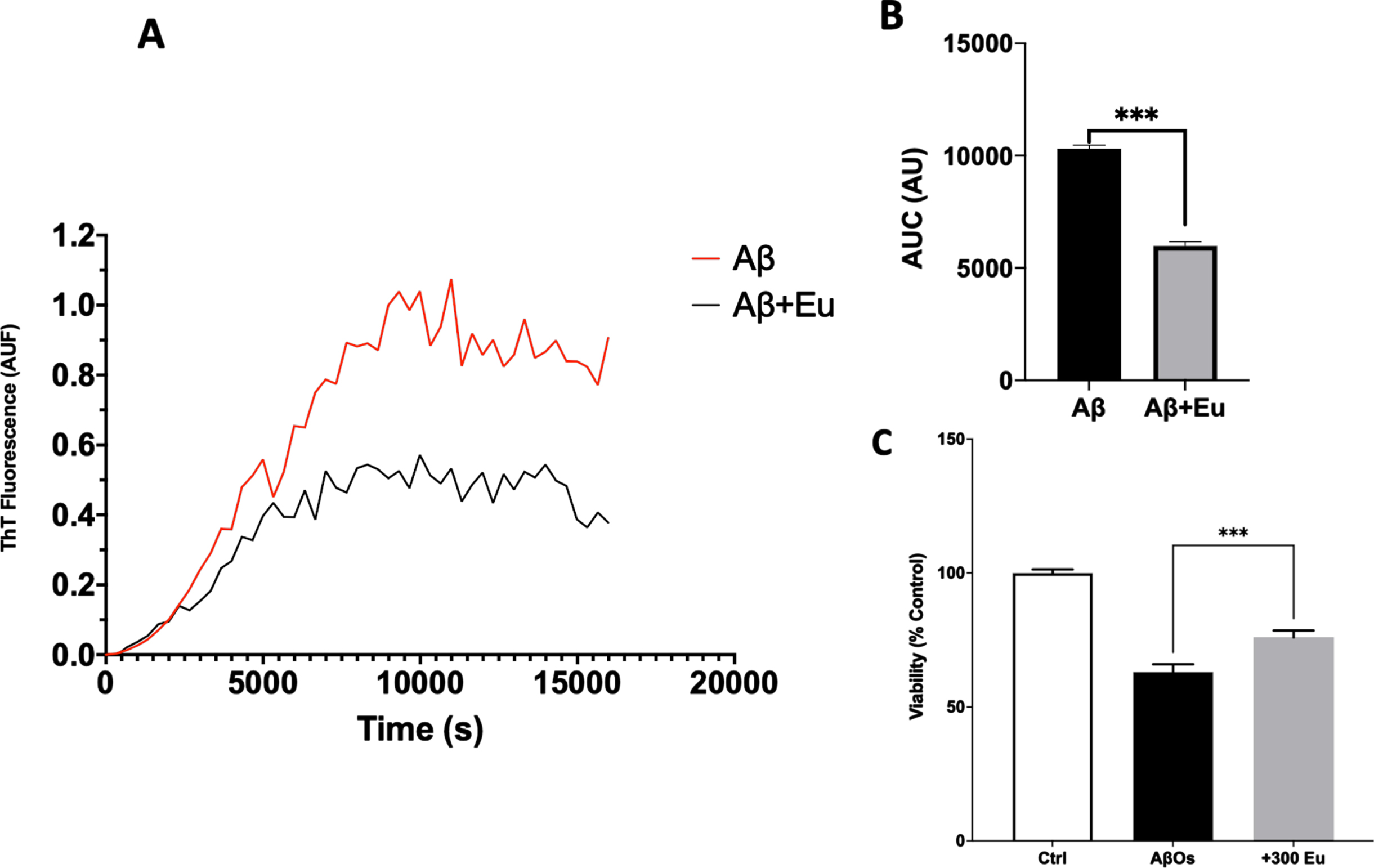
Considering the Eu neuroprotective abilities and synaptic function preservation, we sought to stablish whether Eu influences the aggregation kinetics and presence of Aβ1-40 toxic species [36, 48]. An aggregation assay using Thioflavin T (ThT) was performed. We monitored the fluorescence of ThT in Aβ and Aβ plus 30 or 300 nM of Eu and observed that Eu (30 nM) decreases the maximum fluorescence of Aβ aggregation pattern (Fig. 5A, black line). Also, it was observed that the area under the curve (AUC, Fig. 5B; Aβ: 9860±325.2 AU; Aβ+Eu: 8428±498.6 AU) is correlated with the PC12 viability assay using Aβ aggregated in the presence of Eu (30 mM, Fig. 5C). This shows a correlation between the capacity of Eu to interfere with the aggregation of Aβ and the ability of this lignan to protect PC12 cells from the toxic effects of Aβ. The effect was more pronounced when a higher concentration of Eu (300 nM) was used (Fig. 6A, B), suggesting a concentration-dependent effect (AUC, Fig. 6B; Aβ: 10313±274.7 AU; Aβ+Eu: 6000±297.0 AU). The AβOs generated in the presence of Eu 300 nM showed to be less toxic that those produced in their absence (Fig. 6C). These results showed that Eu has an additional mechanism of action, which promotes neuronal protection and is complementary with the previously observed synaptic effects (structural and functional).
Fig. 5
Eudesmin effects on Aβ aggregation. Aβ1-40 (80μM) aggregation was measured using Thioflavin T (ThT 20μM) evaluating fluorescence every 3 min for 350 min at 37°C, in absence of Eudesmin 30 nM (red) or its presence (black) (A). Area under curve quantification (arbitrary units) shown the effect of Eudesmin over the background fluorescence of ThT (B). Viability quantification evaluated on PC12 cells using MTT to compare response against AβOs or AβOs+Eu (30 nM) (C). Values are mean±SEM, n = 3 using one-way ANOVA and Tukey or Welch’s test. *p < 0.05.
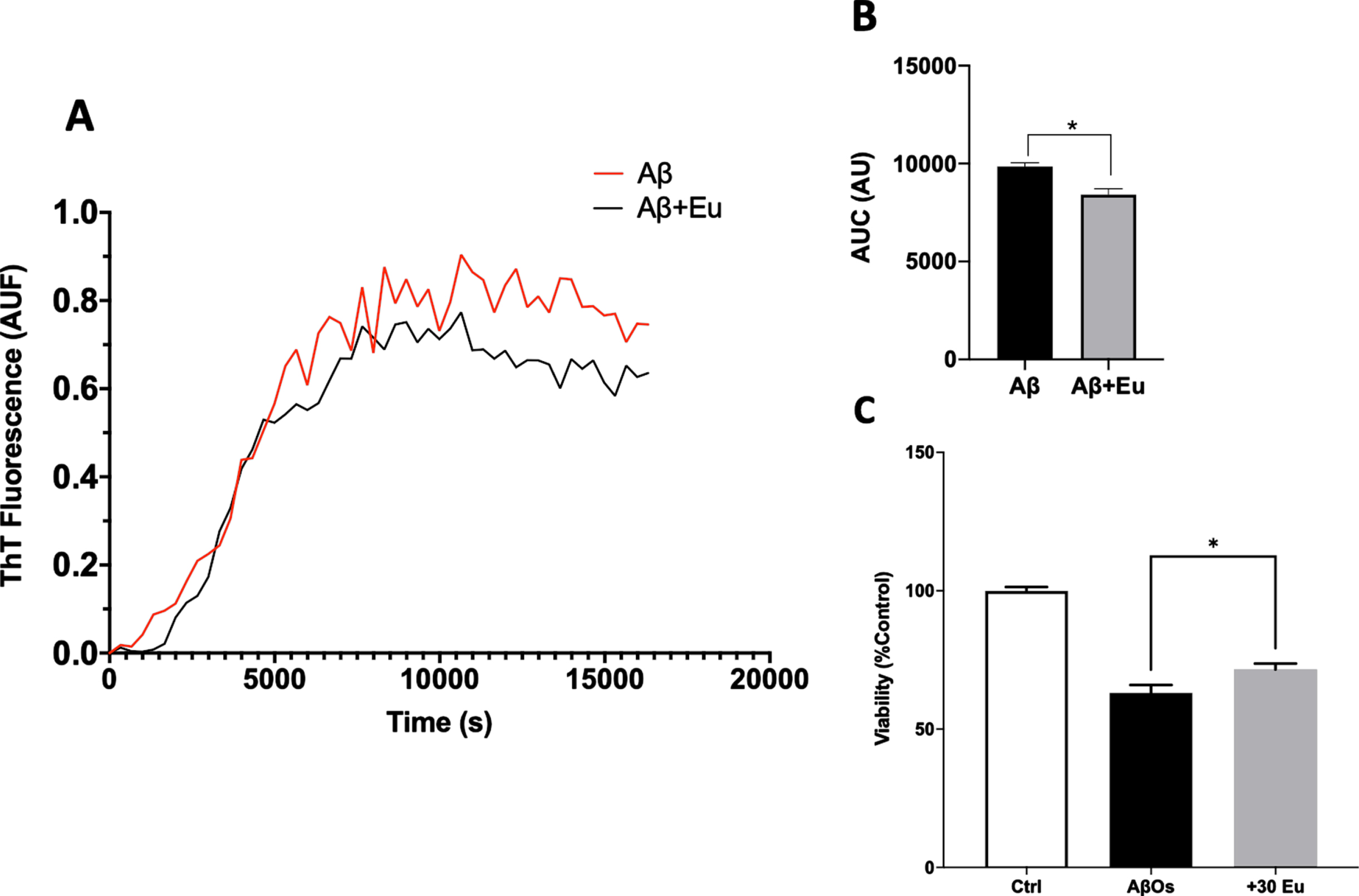
Fig. 6
Eudesmin effects on Aβ aggregation depends on concentration. Aβ1-40 (80μM) aggregation was measured using Thioflavin T (ThT 20μM) evaluating fluorescence every 3 min for 350 min at 37°C, in absence of Eudesmin 300 nM (red) or in presence (black) (A). Area under curve quantification (arbitrary units) shown the effect of Eudesmin over the background fluorescence of ThT (B). Viability quantification evaluated on PC12 cells using MTT to compare response against AβOs or AβOs+Eu (300 nM) (C). Values are mean±SEM, n = 3 using one-way ANOVA and Tukey or Welch’s test. ***p < 0.001.
Eudesmin induces a Aβ-Eu stable complex that could be responsible for reduced AβOs toxicity, in silico evaluation
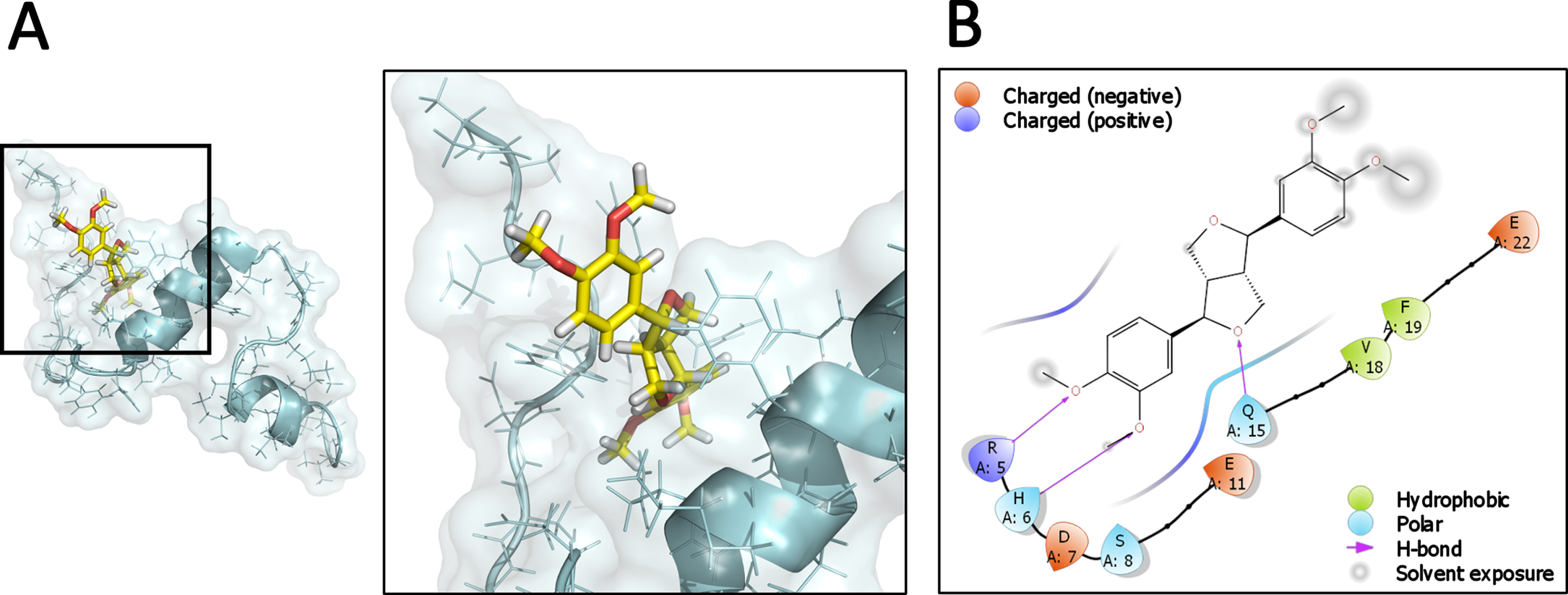
We sought to evaluate the interaction of Eu with Aβ1-40 monomers using in silico techniques. The NMR structure of the Aβ monomer (PDB ID: 1AML) was used for prediction corresponding to Aβ1-40. First, a complex using docking protein-ligand was created (Fig. 7A) in order to evaluate the ability of Eu interaction with Aβ monomer. Eu was able to interact favorably in structural and energetic terms showing a docking score of –2.91 calculated by Glide and a ΔG bind of –48.44 kcal/mol obtained using an MM-GBSA approach calculated with Prime. A more detailed analysis of the Aβ-Eu interaction site showed a region that includes the amino acids 1DAEFRHDS8, 15Q, 18V, and 22E, detecting the formation of 3 hydrogen bonds with the 5R, 6H, and Q15, in a primarily polar region (Fig. 7B). Thus, these results predict a stable and favorable interaction between Eu and the Aβ monomer at the initial state prior to aggregation.
Fig. 7
Predicted interaction of the Aβ1-40 monomer with Eudesmin. Complex between Aβ and Eudesmin generated by docking protein-ligand. Cartoon representation and surface of Aβ1-40 monomer has been colored cyan, while the carbon backbone and oxygen atoms of Eudesmin are shown yellow and red, respectively (A). Schematic representation of the interactions presents at the Aβ-Eu interface in a region defined with a cutoff of 4Å (B).
DISCUSSION
The discovery of natural products displaying neuroprotective and synaptic preservation allows a new strategy for treating or preventing the development of diseases related to cognitive impairment, such as AD [30, 36, 49]. In this work, we uncover the previously unknown neuroprotective properties of Eu, a lignan containing a tetrahydrofurofuran core. It has been reported that other lignan similar to Eu, Sesamin, has showed neuroprotective effects involving anti-inflammatory paths [50, 51]. We isolated Eu from Araucaria araucana, a native conifer from Chile that indigenous people have used as food and traditional medicine [41]. Eu did not show toxicity in the concentration range investigated (1–1000 nM), using a cellular viability assay (Supplementary Figure 1). However, Eu showed neuroprotective effects in vitro, in PC12 cell lines and neuronal primary cultures assays [31, 36, 46, 48]. Remarkably, Eu proved to be rather potent, showing neuroprotective effects at 30 nM (in PC12 cells and neuronal cultures) instead of 1μM, as our previously reports for a natural product such as fomannoxin [36]. Eu also showed beneficial effects on synaptic preservation. The expression of SV2 protein as presynaptic marker was evaluated by immunofluorescence and western blot at the same neuroprotective concentration, and we found that Eu (30 nM) preserves the SV2 protein expression during chronic AβOs treatments (24 h). Similar results have been reported in other in vitro models for AD where SV2 expression was diminished by pore formation induced by Aβ oligomers and characterized by calcium entry inducing an ionic dys-homeostasis [12, 35, 47]. Also, a reduced detection of SV2 protein was observed on AD patients [12]. In our study, we observed a strong neuroprotective effect of Eu by conserving the synaptic structure as well as maintaining synaptic communication. To analyze neuronal transmission, we evaluate the effect of Eu co-treatments on calcium transient’s frequency as an indicator of synaptic function, at the same concentration previously described as neuroprotective (30 nM). Eu does not affect the calcium transient’s frequencies observed under control conditions and prevents the detrimental effect of AβOs on Ca+2 homeostasis. Considering that neurotransmitter release is correlated to intracellular calcium, we expected that Eu preserves the presynaptic proteins and the intracellular calcium transients, similarly to other natural products with neuroprotective activity [52]. Thus, we propose an intracellular mechanism that explains the neuroprotective activity of Eu by presynaptic structure preservation, including SV2 and calcium transients. These data are related to similar compounds previously described by our group, including benzofurans that have been reported with the capacity to interfere with the Aβ aggregation process [31, 35, 36]. Using ThT assays, we evaluated the effects of Eu on aggregation process, and we quantify the area under the curve. Then, the viability response was analyzed using cellular model experiments in the presence of AβOs and co-treated with Eu. At low concentrations of Eu (30 nM), a slight effect on Aβ aggregation was observed, and therefore a minor neuroprotective effect on viability response was detected. However, at high concentrations of Eu (300 nM), an increase in both aggregation and viability response effects was detected. These concentration-related effects of Eu on the AβOs aggregation led us to investigate the Aβ-Eu complex, using in silico techniques. These results indicate the stable formation of complexes where key hydrophobic residues such as phenylalanine and valine are required for the Eu effect on the aggregation of Aβ. Also, we observe three specific hydrogens bonds that can stabilize the Aβ-Eu complex formation. The stable Aβ-Eu complex could be responsible for the reduced toxicity observed on viability assays.
Finally, we believe that Eu represents a novel lead that reduces the Aβ toxicity, opening new research venues for lignans as neuroprotective agents. Future efforts will focus on in vivo experimental approaches in order to gain a deep understanding of the neuroprotective mechanism of Eu.
ACKNOWLEDGMENTS
ANID Fondecyt Regular 1190652 (JCP), ANID MEC80190048 (JCP), ANID Fondecyt Regular 1200908 (JFA), ANID Beca Doctorado nacional (JG) 21200612, Fondecyt iniciacion 11221211 (CFB) and Instituto Antártico Chileno (INACH) RT_16_21 (JCP). In addition, this publication was also supported by Universidad del Bío-Bío, DICREA [2220310 AD/EQ] (RAR).
Authors’ disclosures available online (https://www.j-alz.com/manuscript-disclosures/22-0935r1).
SUPPLEMENTARY MATERIAL
[1] The supplementary material is available in the electronic version of this article: https://dx.doi.org/10.3233/JAD-220935.
REFERENCES
[1] | Selkoe DJ ((2002) ) Alzheimer’s disease is a synaptic failure. Science 298: , 789–791. |
[2] | Saez-Orellana F , Octave JN , Pierrot N ((2020) ) Alzheimer’s disease, a lipid story: Involvement of peroxisome proliferator-activated receptor alpha. Cells 9: , 1215. |
[3] | Hardy J , Selkoe DJ ((2002) ) The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 297: , 353–356. |
[4] | Karran E , Mercken M , De Strooper B ((2011) ) The amyloid cascade hypothesis for Alzheimer’s disease: An appraisal for the development of therapeutics. Nat Rev Drug Discov 10: , 698–712. |
[5] | De Strooper B , Karran E ((2016) ) The cellular phase of Alzheimer’s disease. Cell 164: , 603–615. |
[6] | Hampel H , Vassar R , De Strooper B , Hardy J , Willem M , Singh N , Zhou J , Yan R , Vanmechelen E , De Vos A , Nistico R , Corbo M , Imbimbo BP , Streffer J , Voytyuk I , Timmers M , Tahami Monfared AA , Irizarry M , Albala B , Koyama A , Watanabe N , Kimura T , Yarenis L , Lista S , Kramer L , Vergallo A ((2021) ) The beta-Secretase BACE1 in Alzheimer’s disease. Biol Psychiatry 89: , 745–756. |
[7] | Craft DL , Wein LM , Selkoe DJ ((2002) ) A mathematical model of the impact of novel treatments on the A beta burden in the Alzheimer’s brain, CSF and plasma. Bull Math Biol 64: , 1011–1031. |
[8] | Fuentealba J , Dibarrart AJ , Fuentes-Fuentes MC , Saez-Orellana F , Quinones K , Guzman L , Perez C , Becerra J , Aguayo LG ((2011) ) Synaptic failure and adenosine triphosphate imbalance induced by amyloid-beta aggregates are prevented by blueberry-enriched polyphenols extract. J Neurosci Res 89: , 1499–1508. |
[9] | Dresser L , Hunter P , Yendybayeva F , Hargreaves AL , Howard JAL , Evans GJO , Leake MC , Quinn SD ((2021) ) Amyloid-beta oligomerization monitored by single-molecule stepwise photobleaching. Methods 193: , 80–95. |
[10] | Xu W , Kawarabayashi T , Matsubara E , Deguchi K , Murakami T , Harigaya Y , Ikeda M , Amari M , Kuwano R , Abe K , Shoji M ((2008) ) Plasma antibodies to Abeta40 and Abeta42 in patients with Alzheimer’s disease and normal controls. Brain Res 1219: , 169–179. |
[11] | An JP , Ha TKQ , Kim HW , Ryu B , Kim J , Park J , Lee CH , Oh WK ((2019) ) Eudesmane glycosides from Ambrosia artemisiifolia (common ragweed) as potential neuroprotective agents. J Nat Prod 82: , 1128–1138. |
[12] | Parodi J , Sepulveda FJ , Roa J , Opazo C , Inestrosa NC , Aguayo LG ((2010) ) Beta-amyloid causes depletion of synaptic vesicles leading to neurotransmission failure. J Biol Chem 285: , 2506–2514. |
[13] | Sepulveda FJ , Parodi J , Peoples RW , Opazo C , Aguayo LG ((2010) ) Synaptotoxicity of Alzheimer beta amyloid can be explained by its membrane perforating property. PLoS One 5: , e11820. |
[14] | Demuro A , Parker I , Stutzmann GE ((2010) ) Calcium signaling and amyloid toxicity in Alzheimer disease. J Biol Chem 285: , 12463–12468. |
[15] | Saez-Orellana F , Fuentes-Fuentes MC , Godoy PA , Silva-Grecchi T , Panes JD , Guzman L , Yevenes GE , Gavilan J , Egan TM , Aguayo LG , Fuentealba J ((2018) ) P2X receptor overexpression induced by soluble oligomers of amyloid beta peptide potentiates synaptic failure and neuronal dyshomeostasis in cellular models of Alzheimer’s disease. Neuropharmacology 128: , 366–378. |
[16] | Sepulveda FJ , Fierro H , Fernandez E , Castillo C , Peoples RW , Opazo C , Aguayo LG ((2014) ) Nature of the neurotoxic membrane actions of amyloid-beta on hippocampal neurons in Alzheimer’s disease. Neurobiol Aging 35: , 472–481. |
[17] | Godoy PA , Ramirez-Molina O , Fuentealba J ((2019) ) Exploring the role of P2X receptors in Alzheimer’s disease. Front Pharmacol 10: , 1330. |
[18] | Saez-Orellana F , Godoy PA , Bastidas CY , Silva-Grecchi T , Guzman L , Aguayo LG , Fuentealba J ((2016) ) ATP leakage induces P2XR activation and contributes to acute synaptic excitotoxicity induced by soluble oligomers of beta-amyloid peptide in hippocampal neurons. Neuropharmacology 100: , 116–123. |
[19] | Saez-Orellana F , Godoy PA , Silva-Grecchi T , Barra KM , Fuentealba J ((2015) ) Modulation of the neuronal network activity by P2X receptors and their involvement in neurological disorders. Pharmacol Res 101: , 109–115. |
[20] | Castillo C , Saez-Orellana F , Godoy PA , Fuentealba J ((2022) ) Microglial activation modulated by P2X4R in ischemia and repercussions in Alzheimer’s disease. Front Physiol 13: , 814999. |
[21] | Jeon SY , Byun MS , Yi D , Lee JH , Choe YM , Ko K , Sohn BK , Choi HJ , Lee JY , Lee DY , Group KR ((2019) ) Influence of hypertension on brain amyloid deposition and Alzheimer’s disease signature neurodegeneration. Neurobiol Aging 75: , 62–70. |
[22] | Choi H , Kim C , Song H , Cha MY , Cho HJ , Son SM , Kim HJ , Mook-Jung I ((2019) ) Amyloid beta-induced elevation of O-GlcNAcylated c-Fos promotes neuronal cell death. Aging Cell 18: , e12872. |
[23] | Francis PT , Palmer AM , Snape M , Wilcock GK ((1999) ) The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J Neurol Neurosurg Psychiatry 66: , 137–147. |
[24] | Eckman CB , Eckman EA ((2007) ) An update on the amyloid hypothesis. Neurol Clin 25: , 669-682, vi. |
[25] | Pritchard SM , Dolan PJ , Vitkus A , Johnson GV ((2011) ) The toxicity of tau in Alzheimer disease: Turnover, targets and potential therapeutics. J Cell Mol Med 15: , 1621–1635. |
[26] | Yan MH , Wang X , Zhu X ((2013) ) Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic Biol Med 62: , 90–101. |
[27] | Koola MM ((2020) ) Galantamine-Memantine combination in the treatment of Alzheimer’s disease and beyond. Psychiatry Res 293: , 113409. |
[28] | Mucke HA ((2015) ) The case of galantamine: Repurposing and late blooming of a cholinergic drug. Future Sci OA 1: , FSO73. |
[29] | Metz CN , Pavlov VA ((2021) ) Treating disorders across the lifespan by modulating cholinergic signaling with galantamine. J Neurochem 158: , 1359–1380. |
[30] | Cabrera-Pardo JR , Fuentealba J , Gavilan J , Cajas D , Becerra J , Napiorkowska M ((2019) ) Exploring the multi-target neuroprotective chemical space of benzofuran scaffolds: A new strategy in drug development for Alzheimer’s disease. Front Pharmacol 10: , 1679. |
[31] | Fuentealba J , Saez-Orellana F ((2014) ) Neuroactive alkaloids that modulate the neuronal nicotinic receptor and provide neuroprotection in an Alzheimer’s disease model: The case of Teline monspessulana. Neural Regen Res 9: , 1880–1881. |
[32] | Khanam H , Shamsuzzaman ((2015) ) Bioactive Benzofuran derivatives: A review. Eur J Med Chem 97: , 483–504. |
[33] | Nevagi RJ , Dighe SN , Dighe SN ((2015) ) Biological and medicinal significance of benzofuran. Eur J Med Chem 97: , 561–581. |
[34] | Chand K , Rajeshwari , Hiremathad A , Singh M , Santos MA , Keri RS ((2017) ) A review on antioxidant potential of bioactive heterocycle benzofuran: Natural and synthetic derivatives. Pharmacol Rep 69: , 281–295. |
[35] | Araya JA , Ramirez AE , Figueroa-Aroca D , Sotes GJ , Perez C , Becerra J , Saez-Orellana F , Guzman L , Aguayo LG , Fuentealba J ((2014) ) Modulation of neuronal nicotinic receptor by quinolizidine alkaloids causes neuroprotection on a cellular Alzheimer model. J Alzheimers Dis 42: , 143–155. |
[36] | Gonzalez-Ramirez M , Gavilan J , Silva-Grecchi T , Cajas-Madriaga D , Trivino S , Becerra J , Saez-Orellana F , Perez C , Fuentealba J ((2018) ) A natural benzofuran from the patagonic Aleurodiscus vitellinus fungus has potent neuroprotective properties on a cellular model of amyloid-beta peptide toxicity. J Alzheimers Dis 61: , 1463–1475. |
[37] | Li SL , Wu HC , Hwang TL , Lin CH , Yang SS , Chang HS ((2020) ) Phytochemical investigation and anti-inflammatory activity of the leaves of Machilus japonica var. kusanoi. Molecules 25: , 4149. |
[38] | Chen FY , Yu WW , Huang JW , Huang WM , Bian YT , Shuang PC , Luo YM ((2021) ) Discovery of eudesmane-type sesquiterpenoids with neuroprotective effects from the roots of Chloranthus serratus. Fitoterapia 153: , 104971. |
[39] | Tang JJ , Wang MR , Dong S , Huang LF , He QR , Gao JM ((2021) ) 1,10-Seco-Eudesmane sesquiterpenoids as a new type of anti-neuroinflammatory agents by suppressing TLR4/NF-kappaB/MAPK pathways. Eur J Med Chem 224: , 113713. |
[40] | Wang X , Peng X , Tang C , Zhou S , Ke CQ , Liu Y , Yao S , Ai J , Ye Y ((2021) ) Anti-inflammatory eudesmane sesquiterpenoids from Artemisia hedinii. J Nat Prod 84: , 1626–1637. |
[41] | Bravo-Arrepol G , Torres S , Figueroa F , Pérez C , Paz C , Turchetti G , Tiezzi A , Becerra J ((2020) ) Chemical characterization of lignans from Araucaria araucana a native conifer of Chile and evaluation of their cytotoxicity and antioxidant activities. J Chil Chem Soc 65: , 4953–4957. |
[42] | Zheng QX , Xu ZJ , Sun XF , Gueritte F , Cesario M , Sun HD , Cheng CH , Hao XJ , Zhao Y ((2003) ) Eudesmane derivatives and other sesquiterpenes from Laggera alata. J Nat Prod 66: , 1078–1081. |
[43] | Kim JY , Seo YH , Lee IH , Choi HY , Kwon HC , Choi JH , Lee J , Jang DS ((2020) ) New eudesmane-type sesquiterpene glycosides from the leaves of Aster koraiensis. Plants (Basel) 9: , 1811. |
[44] | Irwin JJ , Sterling T , Mysinger MM , Bolstad ES , Coleman RG ((2012) ) ZINC: A free tool to discover chemistry for biology. J Chem Inf Model 52: , 1757–1768. |
[45] | Trott O , Olson AJ ((2010) ) AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 31: , 455–461. |
[46] | Gavilan J , Mennickent D , Ramirez-Molina O , Trivino S , Perez C , Silva-Grecchi T , Godoy PA , Becerra J , Aguayo LG , Moraga-Cid G , Martin VS , Yevenes GE , Castro PA , Guzman L , Fuentealba J ((2019) ) 17 oxo sparteine and lupanine, obtained from Cytisus scoparius, exert a neuroprotection against soluble oligomers of amyloid-beta toxicity by nicotinic acetylcholine receptors. J Alzheimers Dis 67: , 343–356. |
[47] | Castillo C , Fernandez-Mendivil C , Buendia I , Saavedra P , Meza C , Parra NC , Lopez MG , Toledo JR , Fuentealba J ((2019) ) Neuroprotective effects of EpoL against oxidative stress induced by soluble oligomers of Abeta peptide. Redox Biol 24: , 101187. |
[48] | Fuentealba J , Dibarrart A , Saez-Orellana F , Fuentes-Fuentes MC , Oyanedel CN , Guzman J , Perez C , Becerra J , Aguayo LG ((2012) ) Synaptic silencing and plasma membrane dyshomeostasis induced by amyloid-beta peptide are prevented by Aristotelia chilensis enriched extract. J Alzheimers Dis 31: , 879–889. |
[49] | Wang X , Yu Z , Wang C , Cheng W , Tian X , Huo X , Wang Y , Sun C , Feng L , Xing J , Lan Y , Sun D , Hou Q , Zhang B , Ma X , Zhang B ((2017) ) Alantolactone, a natural sesquiterpene lactone, has potent antitumor activity against glioblastoma by targeting IKKbeta kinase activity and interrupting NF-kappaB/COX-2-mediated signaling cascades. J Exp Clin Cancer Res 36: , 93. |
[50] | Cheng FC , Jinn TR , Hou RC , Tzen JT ((2006) ) Neuroprotective effects of sesamin and sesamolin on gerbil brain in cerebral ischemia. Int J Biomed Sci 2: , 284–288. |
[51] | Ruankham W , Suwanjang W , Wongchitrat P , Prachayasittikul V , Prachayasittikul S , Phopin K ((2021) ) Sesamin and sesamol attenuate H2O2-induced oxidative stress on human neuronal cells via the SIRT1-SIRT3-FOXO3a signaling pathway. Nutr Neurosci 24: , 90–101. |
[52] | Zurita MP , Munoz G , Sepulveda FJ , Gomez P , Castillo C , Burgos CF , Fuentealba J , Opazo C , Aguayo LG ((2013) ) Ibuprofen inhibits the synaptic failure induced by the amyloid-beta peptide in hippocampal neurons. J Alzheimers Dis 35: , 463–473. |


