
Heightened Tameness and Accelerated Handling-Habituation in 3×Tg-AD Mice on a B6;129 Genetic Background
Abstract
Background:
The triple transgenic mouse model of Alzheimer’s disease (3×Tg-AD) has gained popularity in Alzheimer’s research owing to the progressive development of both amyloid-β and tau pathologies in its brain. Prior handling-habituation, a necessary preparation procedure that reduces anxiety and stress in rodents, was seldom described in the literature involving these mice and needs to be addressed.
Objective:
We sought to determine whether 3×Tg-AD mice differ from B6;129 genetic control mice in terms of tameness and prior habituation to handling.
Methods:
We devised hand-staying and hand-boarding assays to evaluate tameness in 3×Tg-AD and B6;129 genetic control mice at 2.5, 7, and 11.5 months of age, representing cognitively pre-symptomatic, early symptomatic and advanced symptomatic stages of the disease, respectively. We monitored the progress of handling-habituation across 8–15 daily handling sessions and assessed the animal behaviors in elevated plus maze.
Results:
We found that 3×Tg-AD mice were markedly tamer than age-matched control mice at the baseline. Whereas it took 2–3 days for 3×Tg-AD mice to reach the criteria for full tameness, it took an average of 7–9 days for young genetic control mice to do so. Prior handling-habituation enhanced risk assessment and coping strategy in mice in elevated plus maze. Completely handling-habituated mice exhibited comparable anxiety indices in the maze regardless of genotype and age.
Conclusion:
These findings collectively point to inherently heightened tameness and accelerated handling-habituation in 3×Tg-AD mice on a B6;129 genetic background. These traits should be carefully considered when behaviors are compared between 3×Tg-AD and the genetic control mice.
INTRODUCTION
Alzheimer’s disease (AD) is the most common cause of dementia in the elderly. Transgenic mouse models of AD are invaluable tools not only in basic research pursuing mechanistic insights into the disease but also in preclinical studies that are aimed to identify biomarkers and therapeutic targets for the diagnosis and treatment of AD [1, 2]. These transgenic models mostly carry either the human APP transgene with mutations that cause familial AD or the human tau transgene with mutations that cause inherited frontotemporal dementia and parkinsonism linked to chromosome 17, or both [3]. Since first introduced in 2003, the triple transgenic mouse model of AD, also known as the 3×Tg-AD mouse, has gained great popularity in the field of AD research owing to the progressive development of both amyloid- β and tau pathologies in its brain [4].
The 3×Tg-AD line carries two independent transgenes—encoding human APP with the Swedish mutation and human 0N4R tau with P301L mutation, respectively—both under the control of Thy1.2 promoter, on a homozygous mutant PS1M146V knock-in background [4]. Overt amyloid-β and tau pathologies are developed in the brain in 3×Tg-AD mice, starting at about 6 and 12 months of age, respectively [5]. Compared to their non-transgenic control strain, 3×Tg-AD mice exhibit impaired spatial reference memory in Morris water maze and Barnes maze beginning at 3-4 months of age or even earlier when there is only intraneuronal accumulation of amyloid-β, but no appreciable amyloid-β plaques or tau pathology [6–11].
Behavioral assessments of 3xTg-AD and the genetic control mice are of significance in both mechanistic and preclinical studies of AD [2, 6, 12]. Interestingly, 3xTg-AD mice seem to exhibit a higher level of anxiety, as evidenced by less time spent in the center of an open field and/or less time in the open arms of an elevated plus maze, than the genetic control mice in some studies, whereas the opposite was observed in others [2, 13–17]. Because handling of mice is generally indispensable in animal behavioral tests and novel handling leads to fear and distress in laboratory rodents [18], it is important to habituate mice to handling prior to behavioral characterization [19]. However, prior handling-habituation was seldom described or discussed in the literature [20]. How long prior handling-habituation is needed for 3xTg-AD and the control mice, a necessary methodological consideration in behavioral tests, remains uncertain. In the present study, we evaluated the tameness and handling-habituation in cognitively pre-symptomatic and symptomatic 3×Tg-AD and age-matched genetic control mice. We found that 3×Tg-AD mice were inherently tamer and significantly easier to be handling-habituated than control mice, whereas prior handling-habituation enhanced risk assessment and coping strategy in mice in the elevated plus maze task.
MATERIALS AND METHODS
Mice
The 3×Tg-AD mice [B6;129-Tg(APPSwe, tauP301L)1Lfa Psen1tm1Mpm/Mmjax] on a B6;129 genetic background (JAX stock #004807) and their approximate genetic control, the B6129SF2/J mice (JAX stock #101045), were purchased from the Jackson Laboratory, and bred and housed in the Laboratory Animal Resource Center at New York State Institute for Basic Research in Developmental Disabilities (IBR) with 12/12 h light/dark cycle and ad libitum access to food and water. This study was carried out in accordance with the PHS Policy on Humane Care and Use of Laboratory animals. All animal procedures were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) at IBR. Homozygous female transgenic and genetic control mice were used for behavioral characterization because female 3×Tg-AD mice are known to show more consistent pathology than their male counterparts [21, 22]. Two cohorts of animals were included, one consisting of two age groups of female mice that began at 2.5 and 7 months of age, respectively, and the other involving both sexes starting at 11.5 months of age. These age groups represented cognitively pre-symptomatic, early symptomatic, and advanced symptomatic stages in 3×Tg-AD mice, respectively [6]. Experimentation on the two cohorts was performed separately. Mice remained undisturbed before their entry into the present study except for weekly cage changes and other husbandry routines. They were transported to a test room and allowed to be acclimated for 1 h before the start of behavioral test sessions.
Assessments of tameness
To compare the baseline level of tameness toward handling between 3×Tg-AD and genetic control mice, we subjected them to hand-staying and hand-boarding tests before the initiation of handling-habituation. To monitor the progress of handling-habituation, we also performed these tests across all daily handling sessions.
For the hand-staying test, the mouse was gently lifted by the base of the tail and placed on the palmar aspect of the experimenter’s hands that formed a scoop and were horizontally positioned at ∼20 cm above the bedding level of a test cage. The latency to fall/jump off the scooped hand was recorded, and then the mouse was returned to its home cage. The cutoff value was set at 20 s.
For the hand-boarding test, the mouse was gently transferred to a test cage that was unoccupied and unscented but otherwise identical to its home cage. After the mouse was allowed to explore the test cage for 30 s, the experimenter extended his scooped hand slowly toward the side of the mouse and tried to position the hand underneath and scoop it up. The untamed non-transgenic mouse generally fled immediately upon the approach attempt. An attempt was considered successful if the mouse could be lifted without jumping away or moving backward and escaping. The cutoff value was set at 20 attempts. The number of successes was recorded, and the success rate calculated for comparison.
Handling-habituation
Mice were subjected to one session of handling-habituation daily for 15 days in 2.5- and 7-month-old mice, and 8 days in 11.5-month-old mice. Prior to the handling-habituation session, tameness was assessed as described above. For each session, mice in a group of four were handled for a total of 5 min. They were placed in one test cage, and then alternately touched, petted, and transferred between test cages. When the 5 min elapsed, mice were returned to their home cage. When a mouse could stay in the scooped hand for no less than 16 s and exhibited a success rate of no less than 80% in hand-boarding assay, we considered that it was completely habituated to handling, i.e., fully tamed.
Elevated plus maze test
The elevated plus maze test, an established assessment of anxiety [23], was conducted as described previously [24] with minor modification. Briefly, the mouse was transferred by hand scoop and placed in the center of the maze facing a closed arm, and allowed to explore the maze for 5 min. The distance traveled and the time spent in each arm and the center area were automatically recorded using the ANY-maze video tracking system (Stoelting Co., Wood Dale, IL, USA). The center and each arm of the maze were defined in the system in such a way that a four-paw criterion was met in considering an entry into an arm. The percentage of time spent in open arms was calculated by dividing the time in both open arms with the total time in all four arms, or alternatively with the total time in the maze. The percentage of distance traveled in open arms was calculated in a similar way. We manually recorded, by using a stopwatch, the time each mouse spent in protected head dipping into either of the two open arms in two scenarios: without entering the open arm and before it eventually entered the open arm as judged by the four-paw criterion. Mice that fell off open arms of the maze during the test were excluded from the statistical analysis of elevated plus maze data unless otherwise specified.
Statistical analysis
Data were analyzed and plotted by using the GraphPad Prism software (GraphPad Software, Inc., San Diego, CA). The Kolmogorov–Smirnov test was used to examine the normality of data distribution. Data were computed as mean±standard error of the mean (SEM). For a 2×2 factorial design, data were analyzed with two-way analysis of variance (ANOVA) followed by Sidak’s multiple comparisons test. For a 2×2×2 factorial design, data were analyzed with three-way ANOVA followed by Tukey’s multiple comparisons test. Data of the time-lapse assessment of tameness were analyzed with repeated-measures two-way or three-way ANOVA, depending on the experimental design. To depict the trend of progress in handling-habituation, the Boltzmann sigmoidal curve or the Gompertz growth curve, whichever better fit the data, was employed. To predict the average days needed for complete tameness in mice, the best curve-fit equation was utilized to calculate the number of days to reach the criteria. Fisher’s exact test was used to compare the percentage of fully tamed mice and to compare the incidence of falling off the elevated plus maze between two groups. p < 0.05 was considered statistically significant.
RESULTS
3×Tg-AD mice exhibit heightened tameness at the baseline
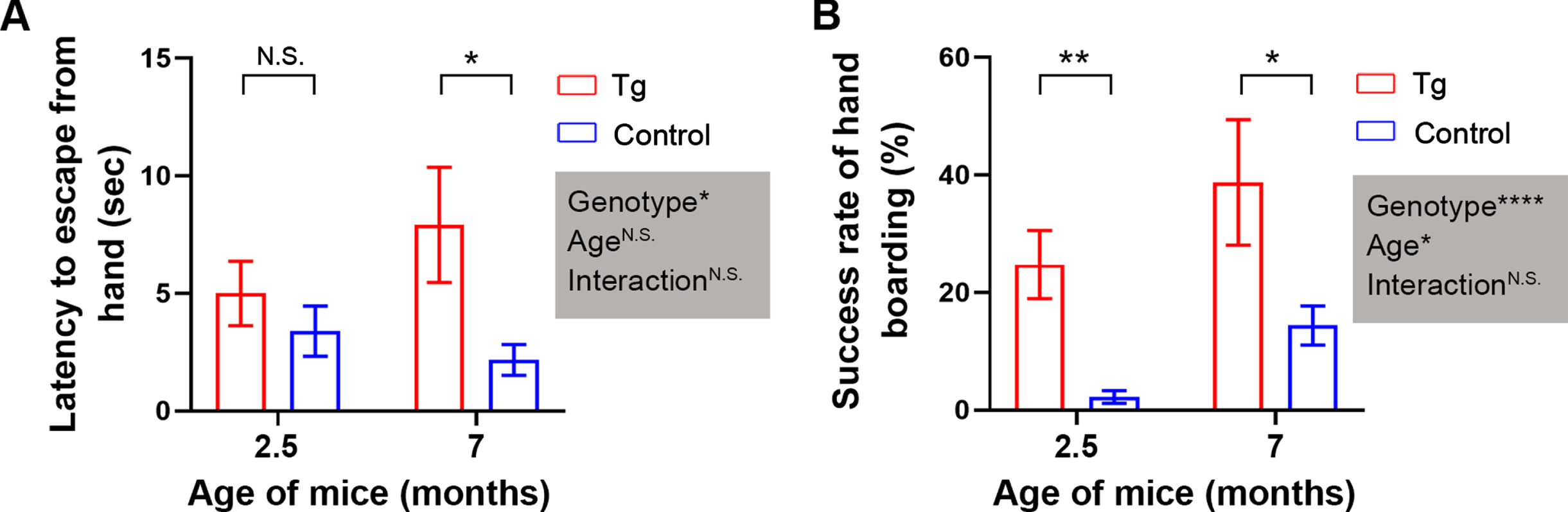
We first characterized the baseline tameness in the first cohort of mice, beginning at 2.5 and 7 months of age. In our assessments of tameness in 3×Tg-AD and B6;129 genetic control mice (Fig. 1), when lifted by the tail and placed on the experimenter’s hand, most of the genetic control mice quickly jumped off the hand, whereas 3×Tg-AD mice remained immobile for a few seconds and then started exploring the hand; some of the 3×Tg-AD mice fell off the hand before the 20 s cutoff time elapsed. When the experimenter tried to lift the unhabituated control mouse by moving one hand toward the side of the mouse, it ran away promptly, and the hand-boarding failed. However, some 3×Tg-AD mice froze in a stereotyped fashion with vocalization upon the approaching attempt and were successfully lifted by scooping, whereas others fled away. The two-way ANOVA results showed a statistically significant main effect for genotype in both assays (Fig. 1A, B) and a main effect for age in hand-boarding success rate (Fig. 1B), with overall longer escape latency and higher boarding success rate in 3×Tg-AD mice than in genetic control mice. Adjusted multiple comparisons test showed statistical significance between 3×Tg-AD and the control mice at both ages in both assays except for the 2.5-month-old mice in hand-staying (Fig. 1A, B). These data suggested that 3×Tg-AD mice appeared inherently tamer than B6;129 genetic control mice, and mice become tamer as they aged regardless of their genotype.
Fig. 1
The baseline level of tameness is markedly higher in 2.5- and 7-month-old 3×Tg-AD mice than in age-matched B6;129 genetic control mice. A) Hand-staying assay. B) Hand-boarding assay. Data are expressed as mean±SEM (n = 12−23 mice/condition) and analyzed with two-way ANOVA. Shaded boxes show main factor effects and the interaction between factors. *p < 0.05, **p < 0.01, ****p < 0.0001. Tg, 3×Tg-AD; control, B6;129 genetic control to 3×Tg-AD; N.S., non-significant. These abbreviations also apply to the other figures unless otherwise specified.
3×Tg-AD mice show accelerated handling-habituation
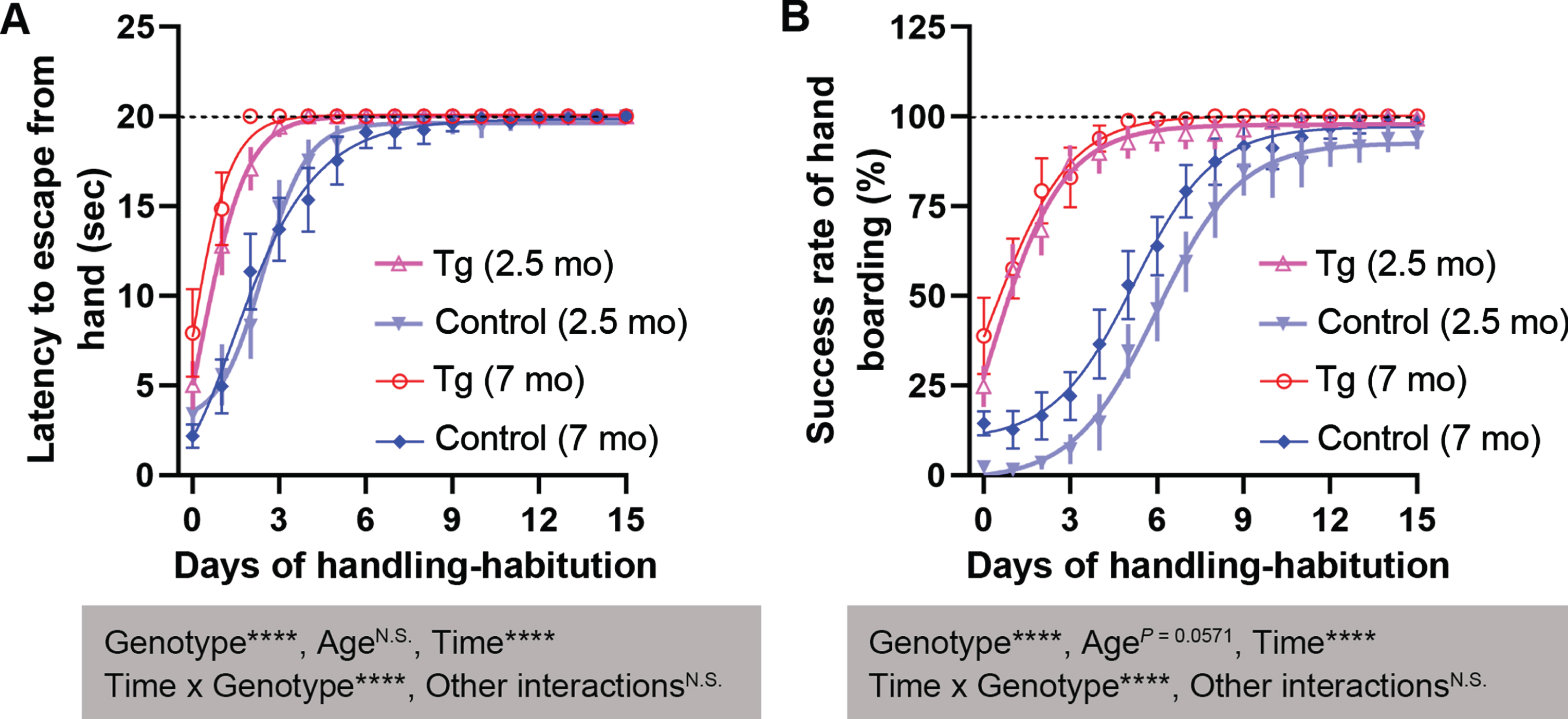
We subjected the 2.5- and 7-month-old mice to daily handling until either the habituation criteria were met or 15 days had elapsed, we plotted the assay results against time (Table 1, Fig. 2). The progress of handling-habituation exhibited curve-fitting by either the Gompertz growth model or the Boltzmann sigmoidal equation (Table 1). In hand-staying and hand-boarding assays, both genotype and time course showed main effects and interaction in repeated-measures ANOVA, with a marked shift of the 3×Tg-AD curves to the left, indicating that it took a markedly shorter time for the 3×Tg-AD mice to be handling-habituated than their control strain (Fig. 2A, B). There was a trend of main effect for age (p = 0.0571) in hand-boarding, as the curve for 2.5-month-old control mice overtly shifted to the right of the curve for their 7-month-old counterparts (Fig. 2B).
Table 1
Curve fit for tameness assays in handling-habituation in 2.5- and 7-month-old mice
| Assay (measure) | Age (mo) | Equation, goodness of fit, and correlation coefficient | |
| 3×Tg-AD | B6;129 genetic control | ||
| Hand-staying (latency to escape) | 2.5 | f(x) =−6.352 + 26.382/(1 + exp((0.2263 − x)/0.8124)) | f(x) = 2.903 + 16.737/(1 + exp((2.424 − x)/0.7605)) |
| R2 = 0.6491, ρ= 0.7612 | R2 = 0.6413, ρ= 0.9588 | ||
| 7 | f(x) = 20.08*(7.697 / 20.08) ∧ exp(−1.345*x) | f(x) = 19.85*(2.110 / 19.85)∧ exp(−0.5995*x) | |
| R2 = 0.5720, ρ= 0.5752 | R2 = 0.6212, ρ= 0.9897 | ||
| Hand-boarding (success rate) | 2.5 | f(x) = 97.73*(27.31 / 97.73)∧ exp(−0.7161*x) | f(x) =−1.392 + 94.052/(1 + exp((5.980 − x)/1.440)) |
| R2 = 0.4939, ρ= 0.9934 | R2 = 0.6556, ρ= 0.9983 | ||
| 7 | f(x) =−11.78 + 111.98/(1 + exp((0.2758 − x)/1.356)) | f(x) = 9.662 + 87.478/(1 + exp((5.196 - x)/1.385)) | |
| R2 = 0.5539, ρ= 0.9354 | R2 = 0.6520, ρ= 0.9912 | ||
mo, months; R2, determination coefficient; ρ, Spearman rank correlation coefficient.
Fig. 2
The 2.5- and 7-month-old 3×Tg-AD mice are handling-habituated more quickly than age-matched B6;129 genetic control mice. A) Hand-staying assay. B) Hand-boarding assay. Data are expressed as mean±SEM (n = 12−23 mice/condition) and analyzed with repeated-measures ANOVA. The Boltzmann sigmoidal and Gompertz growth models are employed for curve fit (see Table 1 for the equations). Shaded boxes show main factor effects and interactions. The dashed line shows the cutoff value (A) or the maximal percentage (B). ****p < 0.0001. mo, months.
We then evaluated the mice individually to see how many days were needed by each mouse to be fully handling-habituated. On average, it took 2–3 days for 3×Tg-AD mice to reach the habituation criteria; by contrast, it took 3–4 and 7–9 days for the control mice to reach the hand-staying and hand-boarding criteria, respectively (Fig. 3A, B). These results are consistent with the predicted days of handling to reach criteria based on calculation using the curve-fit models (Table 2).
Fig. 3
Shorter time is needed for 2.5- and 7-month-old 3×Tg-AD mice to be fully handling-habituated than for age-matched B6;129 genetic control mice. A, B) Bar charts showing days needed to reach the criterion in hand-staying test (A) and in hand-boarding assay (B). Data are expressed as mean±SEM (n = 12−23 mice/condition) and analyzed with two-way ANOVA. Shaded boxes show main factor effects and the interaction between factors. C) Stacked bar charts showing the number of mice that were fully tamed compared to those not fully tamed after 5, 10, and 15 days of handling-habituation. Contingency data were analyzed with Fisher’s exact test. Percentages of completely tamed mice are shown. *p < 0.05, ***p < 0.001, ****p < 0.0001. m.a., months of age.
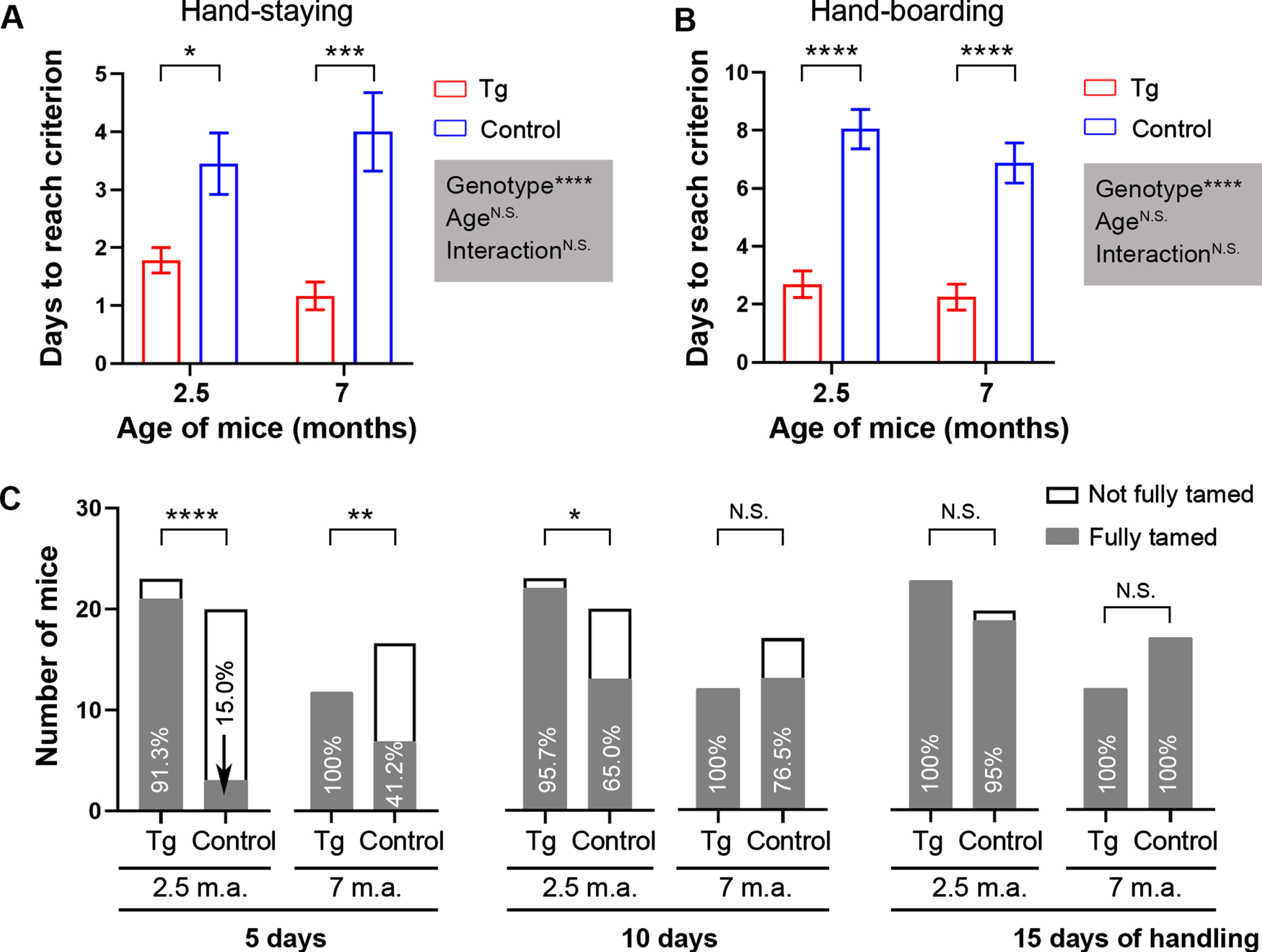
Table 2
Predicted days of handling to reach criteria for full tameness in 2.5- and 7-month-old mice
| Assay | Age (mo) | 3×Tg-AD | B6;129 genetic control | ||
| Number of days | Predicted assay result | Number of days | Predicted assay result | ||
| Hand-staying | 2.5 | 1.62 | 16.01 s | 3.40 | 16.01 s |
| 7 | 1.08 | 16.04 s | 3.91 | 16.01 s | |
| Hand-boarding | 2.5 | 2.59 | 80.05% | 8.66 | 80.00% |
| 7 | 2.33 | 80.02% | 7.16 | 80.08% | |
mo, months; s, seconds.
After 5 days of handling, over 91% of 3×Tg-AD mice were fully tamed, whereas only 15–41.2% of genetic control mice were completely tamed, with a lower percentage for the younger age (Fig. 3C). After 10 days of handling, the percentage for the fully tamed mice of the control strain rose to 65–76.5%, but the 2.5-month-old 3×Tg-AD mice were still markedly tamer than age-matched control mice (Fig. 3C). Two of 37 control mice did not reach the habituation criterion set for hand-boarding by 15 days, despite the fact that no statistical significance was detected between groups after the extended handling-habituation process (Fig. 3C).
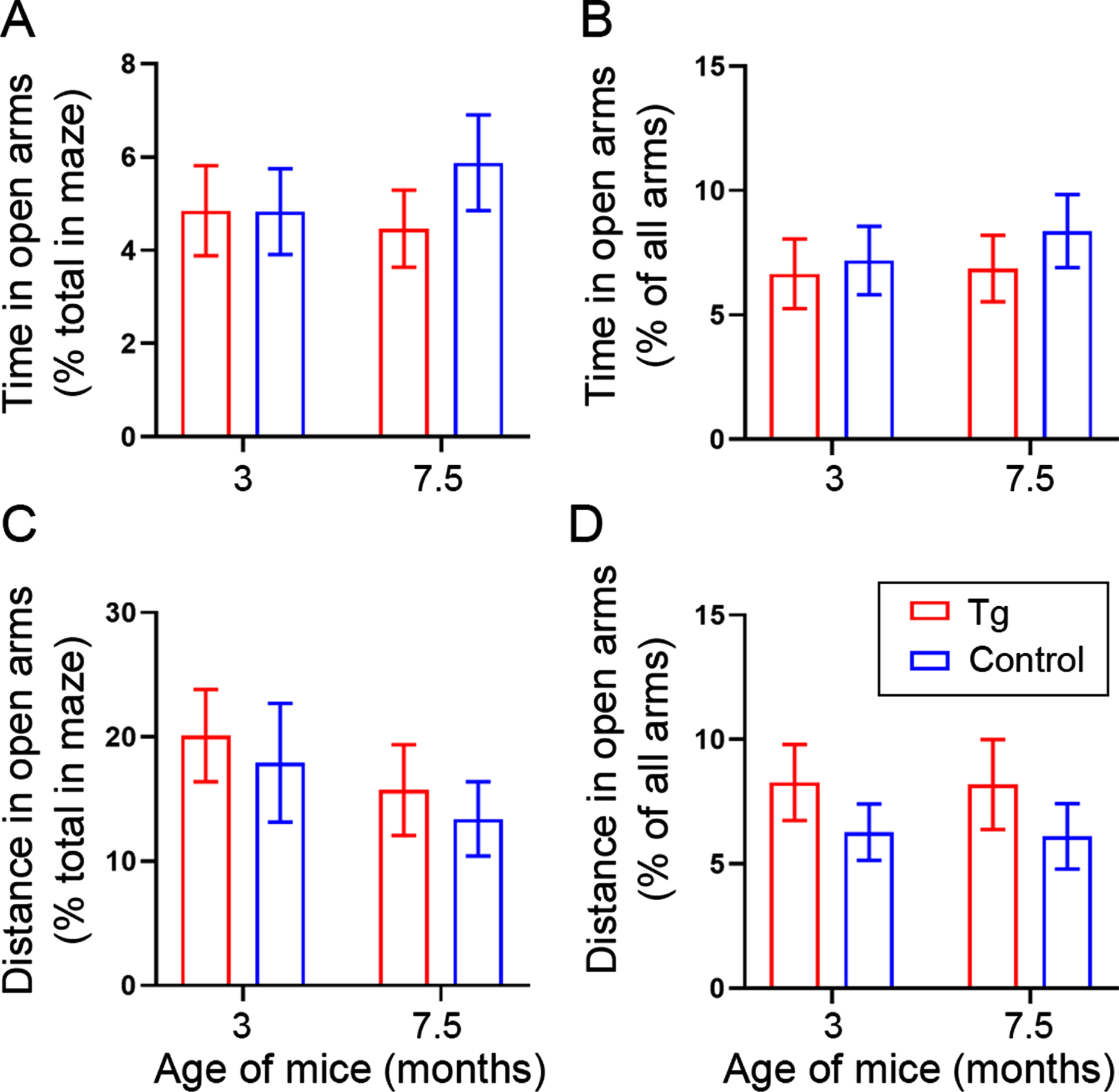
To know whether the 2.5- and 7-month-old 3×Tg-AD mice and their control strain exhibited different levels of anxiety, we subjected the completely handling-habituated mice to elevated plus maze test. We found that the anxiety indices for 3×Tg-AD mice did not statistically differ from those for the age-matched control mice, as evidenced by comparable percentages of time and distance traveled in the open arms (Fig. 4).
Fig. 4
Completely tamed 3×Tg-AD and control mice exhibit comparable anxiety-like behavior in elevated plus maze at the age of 3 and 7.5 months. The percentages were calculated by dividing the time and distance in open arms with total time and total distance in the maze including the center area (A, C) or alternatively with the time and the distance in all four arms (B, D), respectively. Data are expressed as mean±SEM (n = 12−23 mice/condition) and analyzed with two-way ANOVA. No statistically significant effect was detected in the main factors, genotype, or age, nor in the interaction.
Middle-aged 3×Tg-AD mice are tamer than age-matched control mice
To learn whether the inherently heightened tameness in young 3×Tg-AD mice is also present in middle-aged transgenic mice where not only amyloid-β but also tau pathology is abundant in the brain [4], we subjected randomly grouped mice in the second cohort – 3×Tg-AD and control mice of both sexes at 11.5 months of age–to handling-habituation. Two-way ANOVA showed that there was a main effect for genotype but not sex, without statistical significance in interaction, at the baseline in both hand-staying and hand-boarding assays (Fig. 5A, B). Across 8-day handling-habituation sessions, the control female mice exhibited slower progress in hand-staying compared to control males and 3×Tg-AD mice of both sexes, as evidenced by a main effect for time and the interaction between genotype and sex and that among genotype, sex, and time (Fig. 5C). In hand-boarding assay, it took male mice shorter time to reach the criterion for full tameness than the female mice. Control mice started at a lower level of tameness, but they caught up within 2-3 days (Fig. 5D). Overall, both 3×Tg-AD and control mice were fully tamed by 3-4 daily sessions of handling-habituation (Fig. 5C, D).
Fig. 5
Middle-aged 3×Tg-AD mice are tamer than age-matched control mice and prior handling-habituation markedly enhances the performance of these mice in the elevated plus maze task. Mice of each sex for each genotype, at the age of 11.5 months, were randomized into handling-habituated and non-habituated groups. A, B) Baseline assays of hand-staying and hand-boarding, respectively. C, D) Curve fits of the progress of handling-habituation with the Gompertz growth model. E) Stacked bars showing reduced incidence of falling off the elevated plus maze in handling-habituated mice and in 3×Tg-AD mice. F) Interleaved symbols showing increased duration of head dipping into the open arms. Data are expressed as mean±SEM and analyzed with two-way (A, B), repeated-measures three-way (C, D), or three-way (F) ANOVA. n = 8–13 mice/condition in (A-F); n = 5–11 mice/condition in (F) due to exclusion of mice that fell off the maze. Data in (E) is analyzed with Fisher’s exact test. *p < 0.05, **p < 0.01, ****p < 0.0001. The shaded boxes show corresponding ANOVA results, including main effects and interactions. The exact p values are shown when they are closed to 0.05. F, female; G, genotype; H, habituation; M, male; N.S., non-significant; S, sex; T, time.
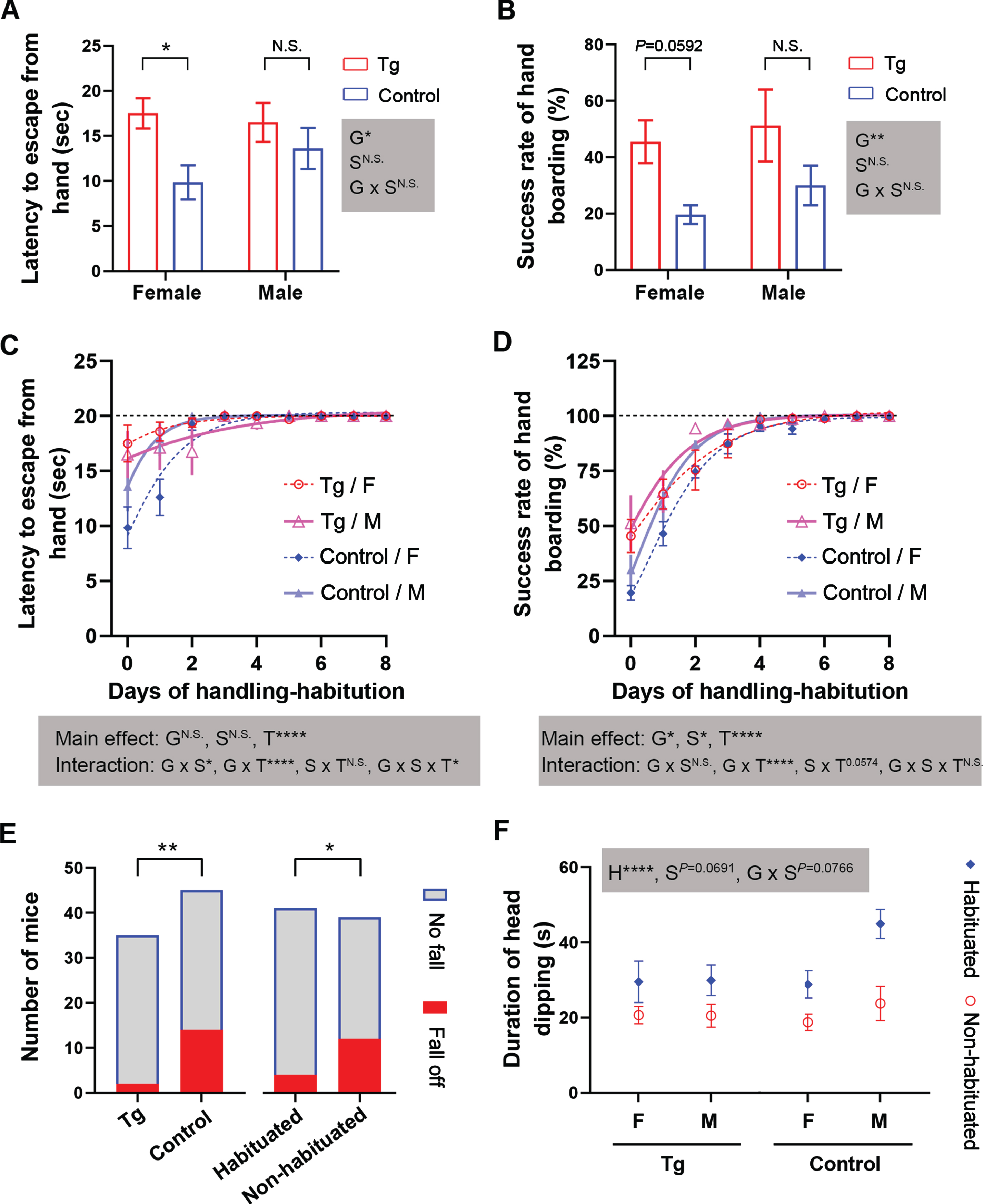
Prior handling-habituation differentially alters the behaviors in elevated plus maze in middle-aged 3×Tg-AD and control mice
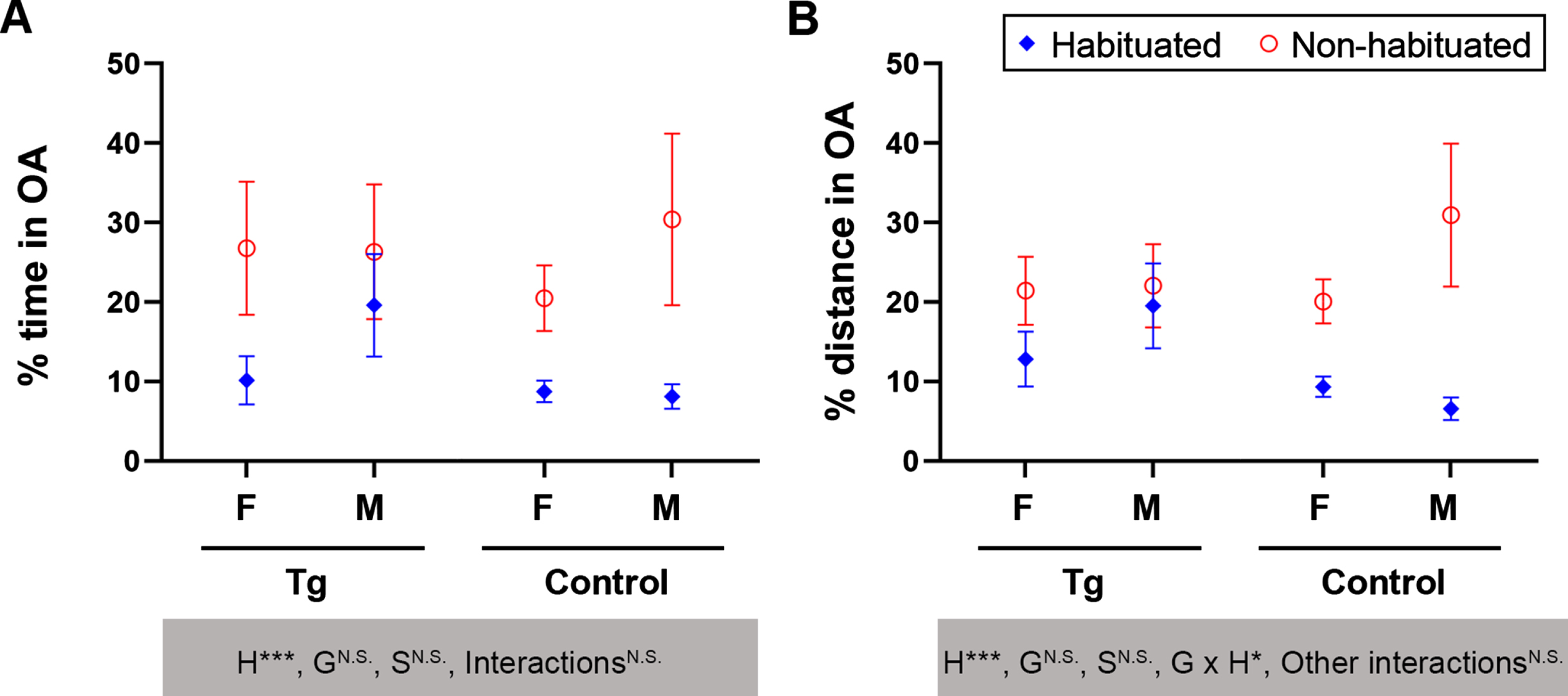
We next subjected the handling-habituated and non-habituated, middle-aged mice to elevated plus maze to explore the influence of prior handling-habituation on the behaviors of mice in this task. We found that unlike fully tamed younger mice in which no falling off the maze was observed, 9.8% of handling-habituated middle-aged mice reversed when exploring the unprotected open arms and fell off the maze. By contrast, 30.8% in non-habituated mice fell (Fig. 5E). The middle-aged 3×Tg-AD mice showed a markedly lower rate of falling off than the control mice (5.7% versus 31.1%, Fig. 5E). Importantly, prior handling-habituated mice exhibited significantly longer duration of protected head dipping into open arms than the non-habituated animals (Fig. 5F). Surprisingly, prior handling-habituated mice showed reduced percentages of time and distance traveled in open arms with diminished within-group variance, especially for control mice of both sexes (Fig. 6A, B).
Fig. 6
Prior handling-habituation markedly reduces the percentages of time (A) and distance traveled (B) in open arms of the elevated plus maze in middle-aged 3×Tg-AD and control mice. The mice were ∼12 months of age when tested in the maze. Data are expressed as mean±SEM (n = 5–11 mice/condition) and analyzed with three-way ANOVA. *p < 0.05, ***p < 0.001. The shaded boxes show corresponding ANOVA results, including main effects and interactions. F, female; G, genotype; H, habituation; M, male; N.S., non-significant; S, sex.
DISCUSSION
Unfamiliar or inappropriate handling can trigger fear and stress in rodents [18]; thus prior habituation to handling is a critical step in preparing rodents for behavioral tests in that it permits a comparable starting point at which animals exhibit minimal anxiety to handling [19]. In the present study, we found that unhandled 3×Tg-AD mice were markedly tamer than the genetic control mice at different stages of cognitive impairment and Alzheimer-like pathologies. The overall progress of handling-habituation for a group of mice was predictable regardless of their genotype or age. In adulthood up to 7 months of age, the transgenic line exhibited significantly accelerated habituation to handling as compared to the genetic control mice; it took significantly longer for B6;129 genetic control mice to be completely habituated to handling than the 3xTg-AD mice. However, the rate of progress in handling-habituation between the two genotypes tended to become comparable at 11.5 months of age despite significantly heightened tameness at the baseline in 3×Tg-AD mice. We also found that fully tamed 3×Tg-AD and B6;129 genetic control mice showed comparable anxiety indices in elevated plus maze and prior handling-habituation reduced fear-like behavior and enhanced the motivation of mice to explore open arms in the maze. These findings have significant implications in handling-habituating 3×Tg-AD and B6;129 genetic control mice for behavioral tests in relevant studies.
Prior habituation to handling is known to help minimize anxiety, stress, and emotional changes in rodents subjected to handling-related procedures [25, 26]. It was shown to significantly reduce the corticosterone level in response to intraperitoneal injection in both adolescent and adult rats [20]. Prior handling-habituation can also promote the interaction of animals with the test apparatus and improve their performance in cognitive tasks [27, 28]. In addition, loss of habituation to handling after memory encoding was negatively correlated to long-term memory retention in a water maze task [29]. Thus, prior handling-habituation can modulate animal behaviors in cognitive assessments in which spontaneous activity and voluntary exploration are involved. Stress and anxiety have a significant neurochemical influence on the amygdala, a brain region that is involved in working memory and the consolidation and retrieval of memory [30, 31]. Therefore, it is possible that prior habituation to handling modulate the amygdala, via which it influences the performance of animals in behavioral tasks.
The number of prior handling-habituation days ranged from 4 to 18 in previous studies, and no criteria were employed for the judgement of complete habituation [9, 14, 26, 28]. In the present study, we devised hand-staying and hand-boarding assays, for the first time to our knowledge, to facilitate the establishment of criteria for complete habituation or full tameness in mice. We proposed to use 80% of the cutoff value, allowing for 20% to cover incidental errors in the assay procedures. We found that it took an average of 7–9 days for 2.5- and 7-month-old B6;129 genetic control mice to reach the criteria for full tameness, as opposed to 2–3 days for 3×Tg-AD mice. When the data of each mouse were analyzed individually, the 2.5-month-old control mice were less tamed than the age-matched 3×Tg-AD mice even after 10 days of handling. These findings suggest that significantly more handling sessions are needed when the B6;129 genetic control strain is included in behavior studies, especially when mice are in young adulthood up to 7 months.
Notably, the 7-month-old 3×Tg-AD mice consistently showed better baseline performance in both hand-staying and hand-boarding assays than age-matched control mice. However, at 2.5 months of age, statistically significant difference at the baseline was detected only in hand-boarding, but not in hand-staying, between transgenic and genetic control mice. These data indicate that hand-boarding is a more stringent assessment of voluntary interaction of mice with the experimenter than is hand-staying. Overall, the 7-month-old mice were tamer than the 2.5-month-old ones at the baseline, regardless of their genotype. This could be the consequence of several months’ longer husbandry routines that included weekly cage changes.
The 3×Tg-AD line was shown to exhibit a higher level of anxiety in elevated plus maze than B6;129 genetic control mice at 1–2 and 3–4 months of age [14]. However, in the present study, we detected no significant difference in anxiety in elevated plus maze between completely tamed 3×Tg-AD and control mice at 2.5 or 7 months of age. This discrepancy may result from different numbers of handling-habituation sessions and possibly different levels of tameness; only five consecutive days of prior handling were employed in the previous study [14]. In another study, 3×Tg-AD mice maintained on a C57BL/6 background at 12–14 months of age did not differ from their non-transgenic control littermates in the percentage of time in open arms of an elevated plus maze [16]. However, in the present study we found heightened baseline tameness in 3×Tg-AD mice at 11.5 months of age. In addition, we detected a greater percentage of control mice that fell off the elevated plus maze than 3×Tg-AD mice at this middle age. These findings suggest that middle-aged 3×Tg-AD mice are tamer, with less intense fear of height, than the B6;129 genetic control mice. The heightened tameness and decreased fear of height in 3×Tg-AD mice could result from overexpression and accumulation of mutant human tau in the amygdala [4], a limbic region known to be critically involved in fear and anxiety [32, 33].
Handling-habituation is known to diminish stress, fear, and anxiety in mice [19], and the elevated plus maze task is an established paradigm to assess anxiety [23]. In this regard, the findings that prior handling-habituation significantly reduced the percentage of time in open arms, a well-recognized measure of anxiety in the elevated plus maze task, are astonishing and opposite to expectation. The only exception is the male 3×Tg-AD mice, in which prior handling-habituation did not show significant effect in percent time or percent distance in open arms. As prior handling-habituation significantly increased the risk assessment behavior, i.e., protected head dipping into open arms [19], the decreased percentages of time and distance in open arms found in the present study could be the consequence of increased perception of risk, which deserves future investigation. Nonetheless, the decreased falling rate indicates that prior handling-habituation markedly reduced fear of height and improved the coping strategy toward acrophobia in these mice.
Cognitive and other behavioral test paradigms that involve handling of mice and that are based on spontaneous activities or non-stringent environmental motivation may be prone to an influence by prior handling-habituation. The questions what dimensions of rodent behaviors are influenced by prior handling-habituation and how remain to be answered. This represents a major limitation of the present study, and these questions deserve future investigations.
In short, we found that adult female 3×Tg-AD mice are markedly tamer and significantly easier to be handling-habituated than B6;129 genetic control mice. These behavioral traits should be carefully considered when the behaviors of 3×Tg-AD mice and the B6;129 genetic control strain are compared.
ACKNOWLEDGMENTS
This work was supported by funds from the New York State Office for People with Developmental Disabilities. We thank Ms. Maureen Marlow of our institute for language editing.
FUNDING
The authors have no funding to report.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
REFERENCES
[1] | Esquerda-Canals G , Montoliu-Gaya L , Guell-Bosch J , Villegas S ((2017) ) Mouse models of Alzheimer’s disease. J Alzheimers Dis 57: , 1171–1183. |
[2] | Blanchard J , Wanka L , Tung YC , Cardenas-Aguayo Mdel C , LaFerla FM , Iqbal K , Grundke-Iqbal I ((2010) ) Pharmacologic reversal of neurogenic and neuroplastic abnormalities and cognitive impairments without affecting Abeta and tau pathologies in 3xTg-AD mice. Acta Neuropathol 120: , 605–621. |
[3] | Myers A , McGonigle P ((2019) ) Overview of transgenic mouse models for Alzheimer’s disease. Curr Protoc Neurosci 89: , e81. |
[4] | Oddo S , Caccamo A , Shepherd JD , Murphy MP , Golde TE , Kayed R , Metherate R , Mattson MP , Akbari Y , LaFerla FM ((2003) ) Triple-transgenic model of Alzheimer’s disease with plaques and tangles: Intracellular Abeta and synaptic dysfunction. Neuron 39: , 409–421. |
[5] | Oddo S , Caccamo A , Kitazawa M , Tseng BP , LaFerla FM ((2003) ) Amyloid deposition precedes tangle formation in a triple transgenic model of Alzheimer’s disease. Neurobiol Aging 24: , 1063–1070. |
[6] | Billings LM , Oddo S , Green KN , McGaugh JL , LaFerla FM ((2005) ) Intraneuronal Abeta causes the onset of early Alzheimer’s disease-related cognitive deficits in transgenic mice. Neuron 45: , 675–688. |
[7] | Stover KR , Campbell MA , Van Winssen CM , Brown RE ((2015) ) Early detection of cognitive deficits in the 3xTg-AD mouse model of Alzheimer’s disease. Behav Brain Res 289: , 29–38. |
[8] | Belfiore R , Rodin A , Ferreira E , Velazquez R , Branca C , Caccamo A , Oddo S ((2019) ) Temporal and regional progression of Alzheimer’s disease-like pathology in 3xTg-AD mice. Aging Cell 18: , e12873. |
[9] | Davis KE , Eacott MJ , Easton A , Gigg J ((2013) ) Episodic-like memory is sensitive to both Alzheimer’s-like pathological accumulation and normal ageing processes in mice. Behav Brain Res 254: , 73–82. |
[10] | Stevens LM , Brown RE ((2015) ) Reference and working memory deficits in the 3xTg-AD mouse between 2 and 15-months of age: A cross-sectional study. Behav Brain Res 278: , 496–505. |
[11] | Baazaoui N , Flory M , Iqbal K ((2017) ) Synaptic compensation as a probable cause of prolonged mild cognitive impairment in Alzheimer’s disease: Implications from a transgenic mouse model of the disease. J Alzheimers Dis 56: , 1385–1401. |
[12] | Billings LM , Green KN , McGaugh JL , LaFerla FM ((2007) ) Learning decreases A beta*56 and tau pathology and ameliorates behavioral decline in 3xTg-AD mice. J Neurosci 27: , 751–761. |
[13] | Baeta-Corral R , Johansson B , Gimenez-Llort L ((2018) ) Long-term treatment with low-dose caffeine worsens BPSD-like profile in 3xTg-AD mice model of Alzheimer’s disease and affects mice with normal aging. Front Pharmacol 9: , 79. |
[14] | Hutton CP , Lemon JA , Sakic B , Rollo CD , Boreham DR , Fahnestock M , Wojtowicz JM , Becker S ((2018) ) Early intervention with a multi-ingredient dietary supplement improves mood and spatial memory in a triple transgenic mouse model of Alzheimer’s disease. J Alzheimers Dis 64: , 835–857. |
[15] | Sterniczuk R , Antle MC , Laferla FM , Dyck RH ((2010) ) Characterization of the 3xTg-AD mouse model of Alzheimer’s disease: Part 2. Behavioral and cognitive changes. Brain Res 1348: , 149–155. |
[16] | Filali M , Lalonde R , Theriault P , Julien C , Calon F , Planel E ((2012) ) Cognitive and non-cognitive behaviors in the triple transgenic mouse model of Alzheimer’s disease expressing mutated APP, PS1, and Mapt (3xTg-AD). Behav Brain Res 234: , 334–342. |
[17] | Hebda-Bauer EK , Simmons TA , Sugg A , Ural E , Stewart JA , Beals JL , Wei Q , Watson SJ , Akil H ((2013) ) 3xTg-AD mice exhibit an activated central stress axis during early-stage pathology. J Alzheimers Dis 33: , 407–422. |
[18] | Balcombe JP , Barnard ND , Sandusky C ((2004) ) Laboratory routines cause animal stress. Contemp Top Lab Anim Sci 43: , 42–51. |
[19] | Hurst JL , West RS ((2010) ) Taming anxiety in laboratory mice. Nat Methods 7: , 825–826. |
[20] | Ristuccia RC , Hernandez M , Wilmouth CE , Spear LP ((2007) ) Differential expression of ethanol-induced hypothermia in adolescent and adult rats induced by pretest familiarization to the handling/injection procedure. Alcohol Clin Exp Res 31: , 575–581. |
[21] | Hirata-Fukae C , Li HF , Hoe HS , Gray AJ , Minami SS , Hamada K , Niikura T , Hua F , Tsukagoshi-Nagai H , Horikoshi-Sakuraba Y , Mughal M , Rebeck GW , LaFerla FM , Mattson MP , Iwata N , Saido TC , Klein WL , Duff KE , Aisen PS , Matsuoka Y ((2008) ) Females exhibit more extensive amyloid, but not tau, pathology in an Alzheimer transgenic model. Brain Res 1216: , 92–103. |
[22] | Vandal M , White PJ , Chevrier G , Tremblay C , St-Amour I , Planel E , Marette A , Calon F ((2015) ) Age-dependent impairment of glucose tolerance in the 3xTg-AD mouse model of Alzheimer’s disease. FASEB J 29: , 4273–4284. |
[23] | Walf AA , Frye CA ((2007) ) The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat Protoc 2: , 322–328. |
[24] | Zhang Y , Wu F , Iqbal K , Gong CX , Hu W , Liu F ((2019) ) Subacute to chronic Alzheimer-like alterations after controlled cortical impact in human tau transgenic mice. Sci Rep 9: , 3789. |
[25] | Boix F , Fernandez Teruel A , Escorihuela RM , Tobena A ((1990) ) Handling-habituation prevents the effects of diazepam and alprazolam on brain serotonin levels in rats. Behav Brain Res 36: , 209–215. |
[26] | Boix F , Fernandez Teruel A , Tobena A ((1988) ) The anxiolytic action of benzodiazepines is not present in handling-habituated rats. Pharmacol Biochem Behav 31: , 541–546. |
[27] | Cibulski L , Wascher CA , Weiss BM , Kotrschal K ((2014) ) Familiarity with the experimenter influences the performance of Common ravens (Corvus corax) and Carrion crows (Corvus corone corone) in cognitive tasks. Behav Processes 103: , 129–137. |
[28] | File SE , Fluck E ((1994) ) Handling alters habituation and response to stimulus change in the holeboard. Pharmacol Biochem Behav 49: , 449–453. |
[29] | Barry H 3rd ((1957) ) Habituation to handling as a factor in retention of maze performance in rats. J Comp Physiol Psychol 50: , 366–367. |
[30] | Roozendaal B , McEwen BS , Chattarji S ((2009) ) Stress, memory and the amygdala. Nat Rev Neurosci 10: , 423–433. |
[31] | Roesler R , Parent MB , LaLumiere RT , McIntyre CK ((2021) ) Amygdala-hippocampal interactions in synaptic plasticity and memory formation. Neurobiol Learn Mem 184: , 107490. |
[32] | Davis M , Rainnie D , Cassell M ((1994) ) Neurotransmission in the rat amygdala related to fear and anxiety. Trends Neurosci 17: , 208–214. |
[33] | Gilpin NW , Herman MA , Roberto M ((2015) ) The central amygdala as an integrative hub for anxiety and alcohol use disorders. Biol Psychiatry 77: , 859–869. |


