
MicroRNA-539 inhibits the progression of Wilms’ Tumor through downregulation of JAG1 and Notch1/3
Abstract
BACKGROUND:
Previous studies demonstrated that miR-539 play an important role in the carcinogenesis of some cancers. The aim of the present study was to determine the role of miR-539 in the pathogenesis of Wilms’ Tumor (WT).
METHODS:
The expression level of miR-539 was measured by qRT-PCR in 42 WT tissues and SK-NEP-1 cell line. Protein expression of genes (E-cadherin, N-cadherin, Vimentin, Notch 1, Notch 3 and JAG1) was assessed by Western blot. The function of miR-539 was investigated in SK-NEP-1 cells by MTT and Transwell assays. The relationship between miR-539 and JAG1 was verified by a dual luciferase assay in SK-NEP-1 cells.
RESULTS:
The expression level of miR-539 was significantly decreased in WT tissues. Downregulation of miR-539 was closely related to NWTS-5 stage, lymph node metastasis and histological type of WT patients. Furthermore, low miR-539 expression was associated with a shorter overall survival rate in WT patients. In vitro, overexpression of miR-539 suppressed proliferation, migration and invasion of SK-NEP-1 cells. In addition, JAG1 was a direct target of miR-539. MiR-539 inhibited the development of WT by inhibiting JAG1-Notch1/3 expressing and blocking EMT.
CONCLUSION:
MiR-539 inhibited the progression of WT through downregulation of JAG1 and Notch1/3.
1.Introduction
Wilms’ tumor (WT) is a malignant embryonal tumor derived from renal blastocyte, accounting for approximately 87% of childhood renal tumors [12]. The peak age of WT is 3 to 4 years old, and 80% of patients suffer WT before the age of 5 [29]. This tumor was first described by Rance in 1814. Max Wilms further described its characteristics in 1899, thus it was named Wilms’ tumor [9]. In addition, there are many treatments for WT. First, all stages of the WT should be surgically removed. Moreover, WT is sensitive to radiotherapy, and postoperative radiotherapy can improve the efficacy [10]. Furthermore, preoperative chemotherapy can shrink the size of the tumor, reduce the risk of surgery, and improve the rate of complete resection [11]. Besides that, the prognosis of WT depends on histologic type, tumor stage, patient age, and biological characteristics [13]. Most WT patients have a good prognosis. After systematic treatment, the 4-year survival rate is approximately 54.8%–90% in WT patients at any stage [26]. To make matters worse, no special precautions were taken for WT. Therefore, early detection and treatment are important to improve the prognosis of WT patients.
Recently, the potential role of microRNAs (miRNAs) in the diagnosis and treatment of cancer has been reported. Many miRNAs have been identified as oncogenes or tumor suppressors involved in the regulation of the tumorigenesis, including WT. For instance, upregulation of miR-190b promoted cell invasion, migration and inhibited cell apoptosis by inhibiting PTEN expression in WT [1]. Cui et al. had been found the prognostic effect of miR-21 in WT patients [6]. Moreover, miR-539 has been found to exhibit different roles in human disease and cancer. On the one hand, miR-539 was upregulated in failing heart [23] and promoted osteoblast proliferation, differentiation and osteoclast apoptosis [33]. On the other hand, Mirghasemi et al. reported that downregulation of miR-539 was associated with unfavorable prognosis in patients with osteosarcoma [22]. It was also found that miR-539 inhibited glioma cell proliferation and invasion by targeting DIXDC1 [25]. However, there is no study on the role of miR-539 in regulating the progression of WT.
As an oncogene, high expression of Jagged-1 (JAG1) has been shown to be associated with the progression and poor prognosis of cancers [27]. Previous studies have also shown that JAG1 expression may be regulated by several miRNA, such as miR-26a [20] and miR-524 [5]. In addition, JAG1 has been reported to be involved in the development of human cancer through the Notch signaling pathway [31]. According to previous reports, Notch signaling pathway is a double-edged sword in cancer [28]. The Notch signaling pathway includes four cell surface transmembrane receptors (Notch 1–4) and five ligands (Jagged-1, Jagged-2, Delta-like 1, Delta-like 3 and Delta-like 4) in mammals [21]. It has been found that abnormal Notch signaling pathway regulates kidney disease [2]. However, the interaction between the miR-539 and JAG1-Notch1/3 signaling pathway remains elusive in WT.
In this study, the function of miR-539 was investigated in WT. Abnormal expression of miR-539 was observed in WT tissues. The interaction between the miR-539 and JAG1-Notch1/3 signaling pathway was also analyzed in WT. We hope that these findings will provide new therapeutic targets for the treatment of WT.
2.Materials and methods
2.1Clinical tissues
Forty-two WT tissues and adjacent tissues were obtained from The Affiliated Yantai Yuhuangding Hospital of Qingdao University after receiving the written informed consent. All patients did not receive any treatment before surgery. Human tissue was frozen in liquid nitrogen and stored in a
2.2Cell cultures
The human WT cell line SK-NEP-1 was purchased from the American Type Culture Collection (ATCC, Rockville, MD, USA), which was selected to investigate the role of miRNA in WT in previous studies [1, 7, 17]. SK-NEP-1 cells were then seeded in RPMI-1640 with 10% fetal bovine serum (FBS) and incubated with 5% CO
2.3Cell transfection
The miR-539 mimic and inhibitor, JAG1 plasmid were purchased from RiboBio (Guangzhou, China) and then were transferred into SK-NEP-1 cells with Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufactures’ protocols.
2.4RNA extraction and quantitative real time polymerase chain reaction (qRT-PCR)
Total RNA containing miRNA was extracted using TRIzol reagent (Invitrogen, USA) to quantify the expression of miR-539 in WT tissues. The synthesis of cDNA was performed by reverse transcription using the TaqMan
2.5Luciferase activity assay
The wild or mutant type of 3’-UTR of JAG1 was inserted into the pGL3 luciferase vector (Promega, Madison) for luciferase reporter experiments. The 3’-UTR of wild or mutant JAG1 and miR-539 mimic were then transfected into SK-NEP-1 cells. Subsequently, dual luciferase assay (Promega, Madison) was used to analyze luciferase activity.
2.6MTT assay for cell proliferation
Cell proliferation was assessed using MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide) assay. Cells (2
2.7Transwell assay
Transwell chambers (8-
2.8Western blot analysis
Protein samples were obtained using RIPA lysis buffer. Proteins were separated by 10% SDS-PAGE and incubated with 5% skim milk in PVDF membranes at room temperature. Next, we plated the membranes at 4
Table 1
Relationship between miR-539 expression and their clinic-pathological characteristics of Wilms’ tumor patients
| Characteristics | Cases | miR-539 | ||
|---|---|---|---|---|
| High | Low | |||
| Age (months) | 0.324 | |||
| | 20 | 9 | 11 | |
| | 22 | 8 | 14 | |
| Gender | 0.206 | |||
| Male | 18 | 8 | 10 | |
| Female | 24 | 9 | 15 | |
| NWTS-5 stage | 0.041* | |||
| I | 30 | 11 | 19 | |
| III | 12 | 4 | 8 | |
| Histological type | 0.0201* | |||
| Favorable (FH) | 32 | 12 | 20 | |
| Unfavorable (UH) | 10 | 4 | 6 | |
| Lymph node metastasis | 0.049* | |||
| No | 28 | 11 | 17 | |
| Yes | 14 | 6 | 8 | |
Statistical analyses were performed by the
Figure 1.
Downregulation of miR-539 was identified in WT tissues.(A) The expressions of miR-539 in WT tissues detected via qRT-PCR. (B) Low miR-539 expression was correlated with shorter overall survival of WT patients. *
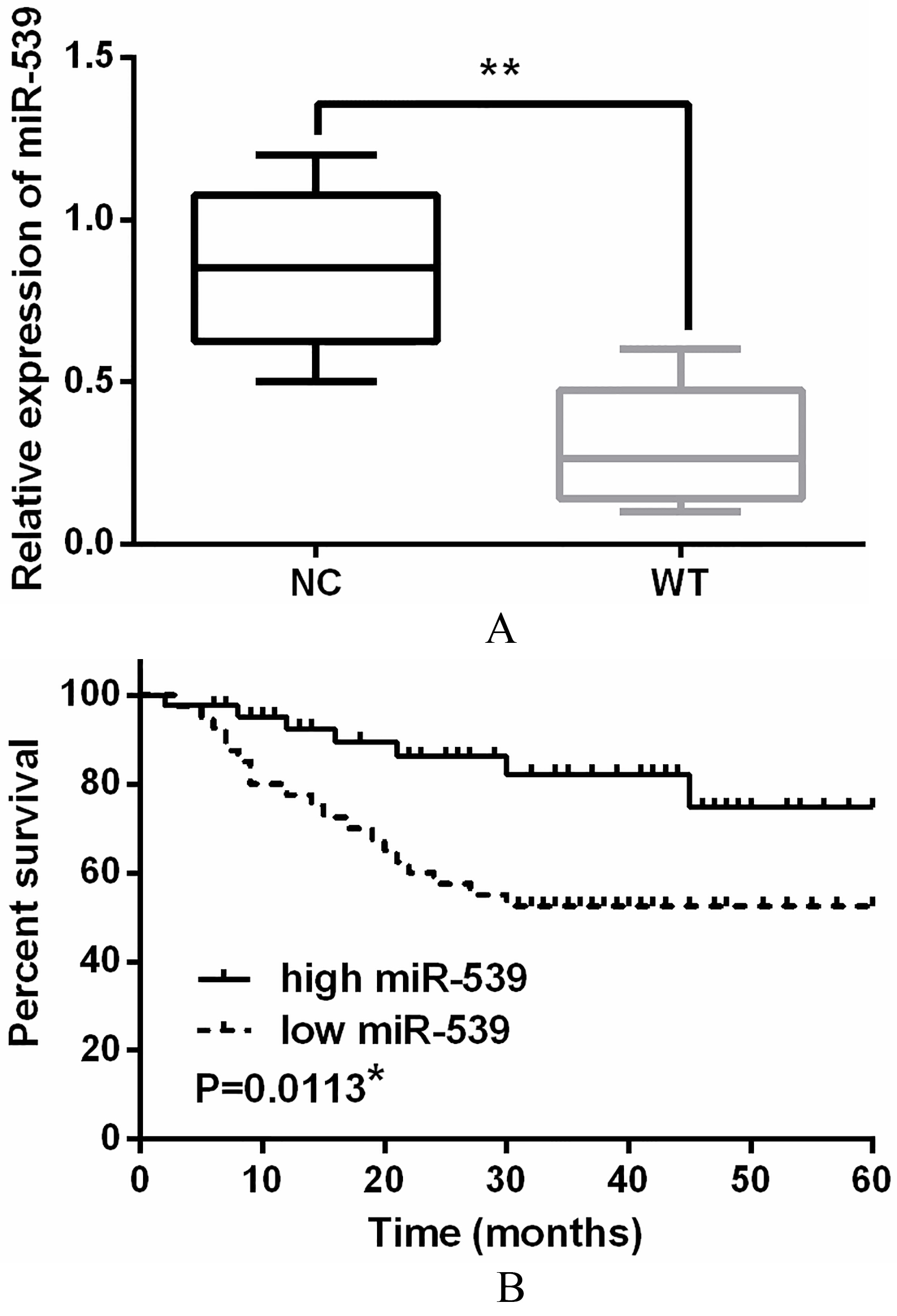
Figure 2.
Overexpression of miR-539 suppressed the proliferation, migration and invasion of WT cells. (A) The miR-539 expression in SK-NEP-1 cell lines. (B) The miR-539 expressions were examined in SK-NEP-1 cells containing miR-539 mimics or inhibitor via qRT-PCR. (C, D) The cell proliferation was measured in cells containing miR-539 mimics or inhibitor via MTT assay. (E, F) Cell migration and invasion analysis in SK-NEP-1 cells containing miR-539 mimics or inhibitor was examined by Transwell assay. *
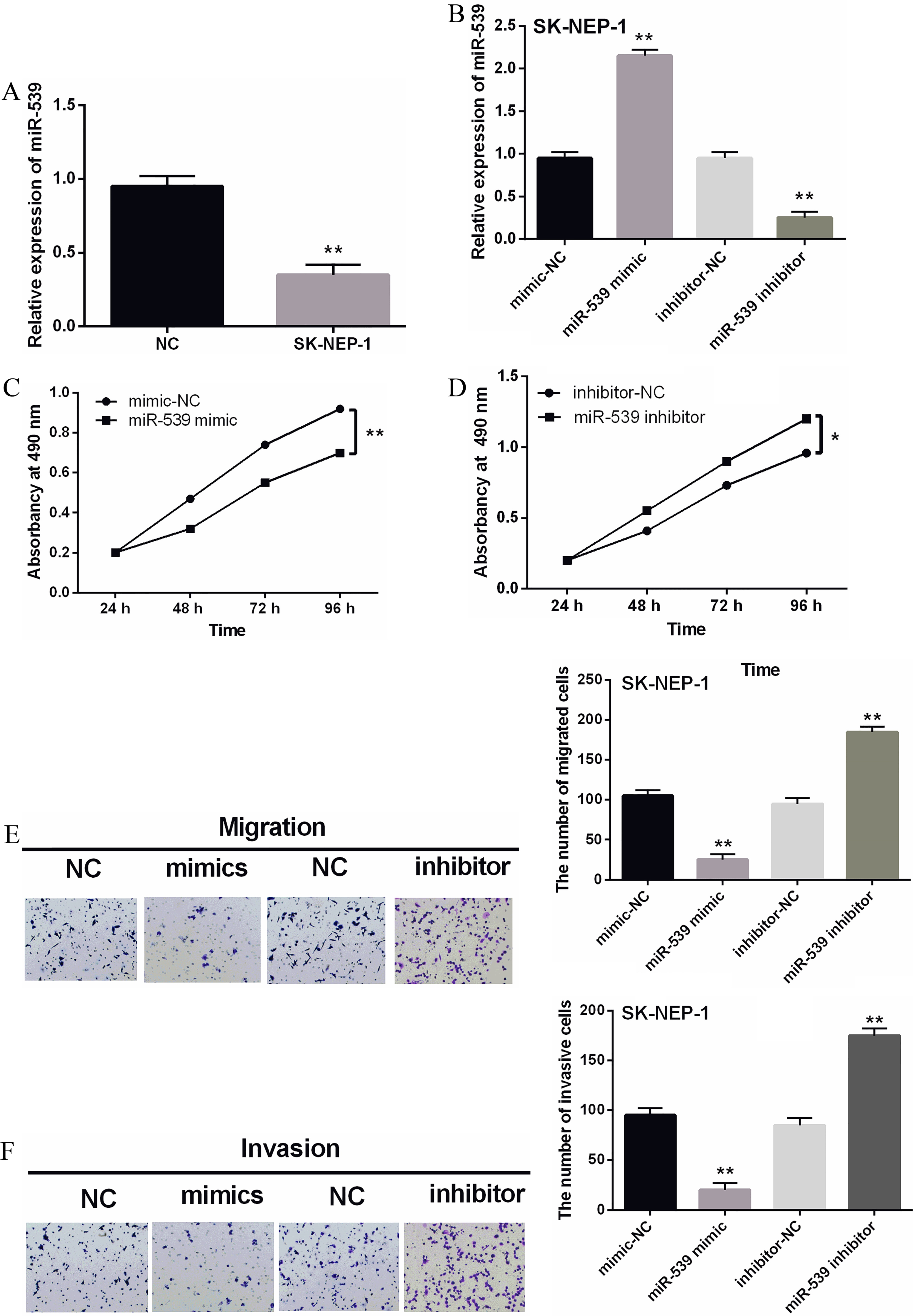
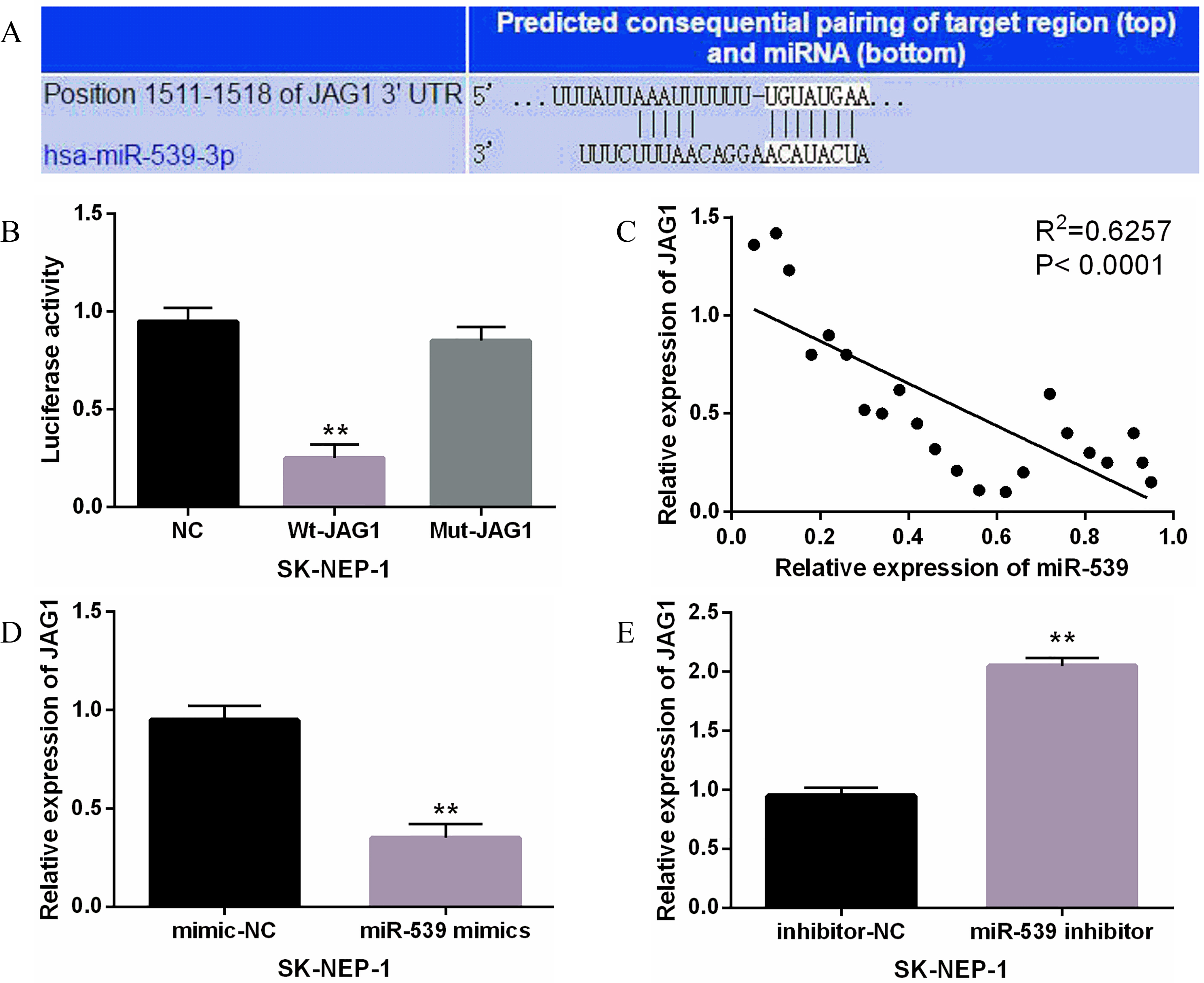
Figure 3.
JAG1 was a direct target of miR-539. (A) JAG1 had binding sites with miR-539. (B) Luciferase reporter assay (C) MiR-539 had negative correlation with JAG1. (D, E) The expression of JAG1 were observed in SK-NEP-1 cells containing miR-539 mimics or inhibitor **
2.9Statistical analysis
Data were shown as mean
3.Results
3.1Downregulation of miR-539 was identified in WT tissues
The expression of miR-539 was detected in WT tissues by qRT-PCR assay. The expression of miR-539 in WT tissues was approximately half of that in normal tissues, indicating that miR-539 was downregulated in WT tissues (Fig. 1A). Moreover, the aberrant expression of miR-539 was closely associated with NWTS-5 stage (
3.2Overexpression of miR-539 suppressed the proliferation, migration and invasion of WT cells
The expression of miR-539 was observed in the SK-NEP-1 cell line. Downregulation of miR-539 was also identified in SK-NEP-1 cells (Fig. 2A). The miR-539 mimics or inhibitor was then transfected into SK-NEP-1 cells to investigate its effect in WT. As shown in Fig. 2B, the expression of miR-539 was enhanced by its mimics and reduced by miR-539 inhibitor. Next, the function of miR-539 was investigated by MTT and Transwell assays. Overexpression of miR-539 was found to suppress proliferation of SK-NEP-1 cells (Fig. 2C). In contrast, silencing of miR-539 promoted cell proliferation in WT (Fig. 2D). Furthermore, up-regulation of miR-539 inhibited migration of SK-NEP-1 cells, while knockdown of miR-539 promoted migration of SK-NEP-1 cells (Fig. 2E). The same results were also identified for cell invasion in SK-NEP-1 cells with miR-539 mimics or inhibitor (Fig. 2F). In conclusion, miR-539 plays an inhibitory role in WT.
Figure 4.
MiR-539 inhibited the development of WT through targeting JAG1. (A) The expression of JAG1 in WT tissues was detected. (B) High JAG1 expression was correlated with shorter overall survival of WT patients. (C) The JAG1 expression was measured in SK-NEP-1 cells with JAG1 vector and miR-539. (D) The cell proliferation was measured in SK-NEP-1 cells with JAG1 vector and miR-539 via MTT. (E, F) The cell migration and invasion in SK-NEP-1 cells with JAG1 vector and miR-539 was measured by Transwell assay. **
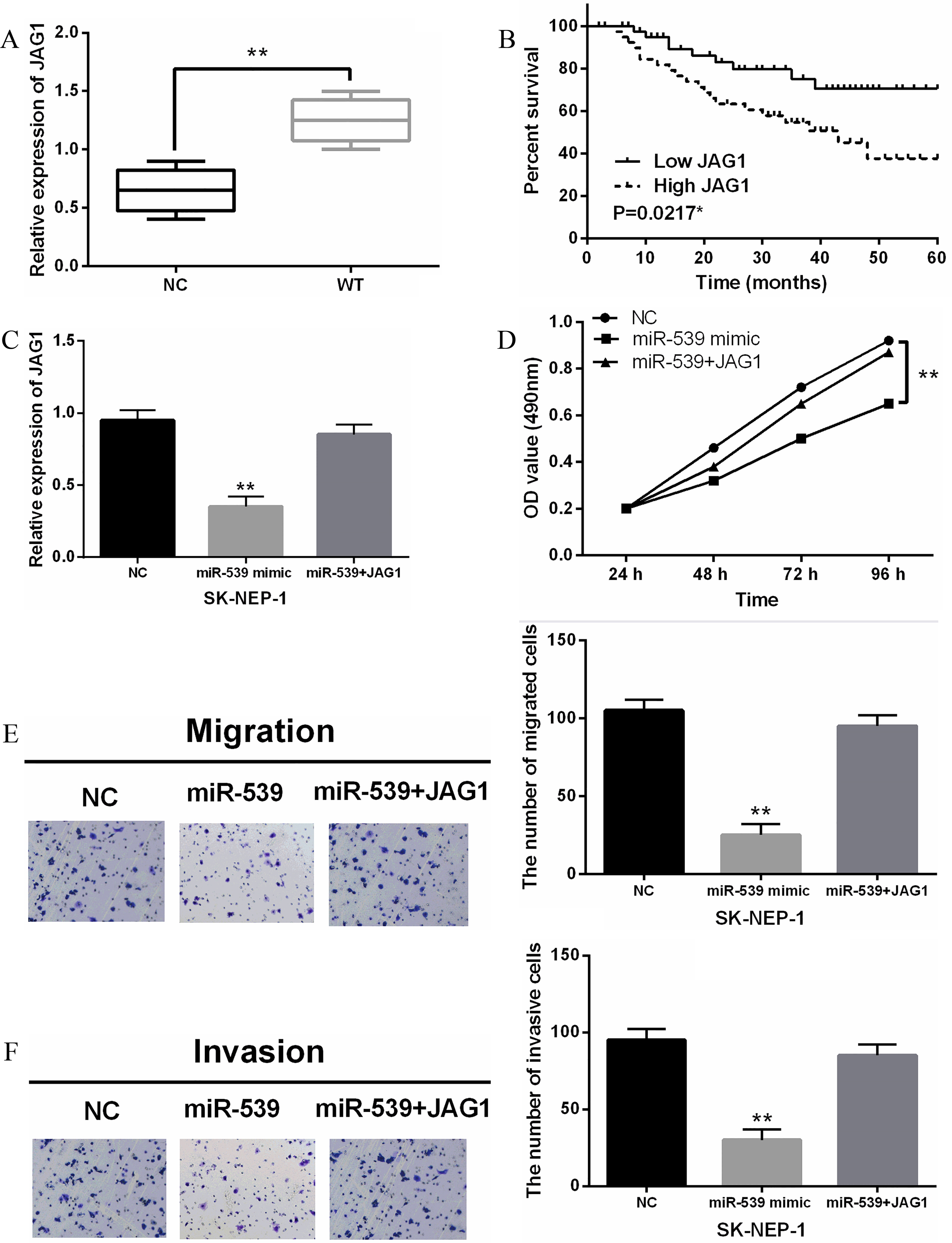
Figure 5.
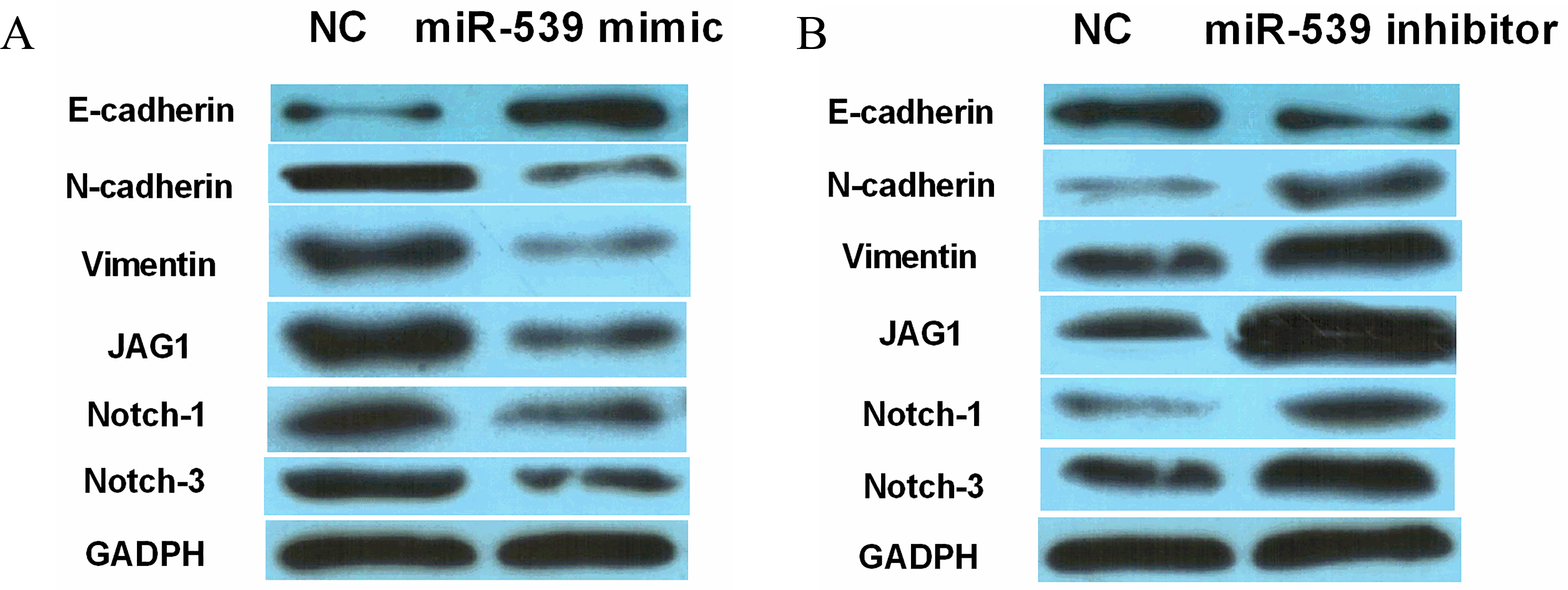
MiR-539 inhibited EMT and Notch1/3 expression in WT. (A, B) The protein expression of E-cadherin, N-cadherin, Vimentin, Notch-1 and Notch-3 in SK-NEP-1 cells contained miR-539 mimics or inhibitor.
3.3JAG1 was a direct target of miR-539
Further, JAG1 was found to have a binding site for miR-539, which was predicted by TargetScan (http://www.targetscan.org/) (Fig. 3A). It suggests that miR-539 may target JAG1. To confirm this prediction, luciferase reporter assay was performed. We found that miR-539 mimics inhibited the luciferase activity of Wt-JAG1. However, the luciferase activity of Mut-JAG1 was not affected by miR-539 mimics (Fig. 3B). Moreover, a negative correlation between miR-539 and JAG1 was identified in WT tissues (
3.4MiR-539 inhibited the development of WT through targeting JAG1
To explore the role of JAG1, we first examined the abnormal expression of JAG1 in WT tissues. The expression of JAG1 was significantly increased in WT tissues compared with normal tissues (Fig. 4A). Moreover, we also found that upregulation of JAG1 predicted worse prognosis in patients with WT (
3.5MiR-539 inhibited EMT and suppressed Notch1/3 expression in WT
Finally, the effect of miR-539 on EMT and Notch1/3 expression was explored to further explain the regulatory mechanism of miR-539 in WT. The results showed that upregulation of miR-539 promoted E-cadherin expression and inhibited N-cadherin and Vimentin expressions in SK-NEP-1 cells, indicating that overexpression of miR-539 inhibited EMT in WT (Fig. 5A). Oppositely, knockdown of miR-539 was found to promote EMT of WT (Fig. 5B). Besides that, JAG1 is a ligand of Notch pathway, which includes other receptors Notch-1, Notch-2 and Notch-3. And we investigated whether the Notch pathway is involved in the development of WT. Western blot analysis showed that the expressions of Notch-1 and Notch-3 were reduced by the miR-539 mimics (Fig. 5A), while the miR-539 inhibitor increased their expressions in SK-NEP-1 cells (Fig. 5B). Based on these results, miR-539 inhibited EMT and suppressed Notch1/3 expression in WT, suggesting that the Notch pathway may be involved in the development of WT.
4.Discussion
As a malignant renal tumor, WT often occurs in children [3]. Previous studies have shown that the progression and development of WT should be mediated by several miRNAs. Liu et al. proposed that miR-19b promoted proliferation and migration of WT cells via the PTEN/PI3K/AKT signaling pathway [17]. Inversely, miR-613 attenuated proliferation, migration and invasion of WT cells by targeting FRS2 [30]. In the present study, it was also found that overexpression of miR-539 inhibited proliferation, migration and invasion of WT cells. Furthermore, expression of miR-539 was reduced in WT tissues and abnormal miR-539 expression was associated with NWTS-5 stage, lymph node metastasis and histological type. In addition, downregulation of miR-539 was related to poor prognosis in WT patients. All of these findings demonstrate that miR-539 acts as a suppressor gene in WT.
It is well known that miR-539 exhibits an inhibitory effect in different human cancers by modulating some target genes. For example, miR-539 suppressed osteosarcoma cell invasion and migration by targeting MMP-8 [16], which was similar to our results in WT. Additionally, miR-539 was found to be downregulated in breast cancer and used as a tumor suppressor [14]. Moreover, miR-539 suppressed cell proliferation and induced cell apoptosis in renal cell carcinoma by targeting HMGA2 [32]. These previous studies strongly supported our conclusions in this study. Further, JAG1 was shown to be a direct target of miR-539 and a negative correlation between miR-539 and JAG1 was identified in WT tissues. In particular, miR-539 inhibited the development of WT through targeting JAG1.
As shown in previous studies, miRNAs inhibit the expressions of many target genes by binding to their 3’-UTR. In this study, identifying downstream target genes of miR-539 is an important task for us. JAG1 has been identified as a target gene in some cancers. For instance, miR-186 inhibited cell proliferation in multiple myeloma by inhibiting JAG1 [19], which was consistent with our results. Besides that, miR-539 inhibited EMT and suppressed the expressions of Notch1/3 in WT. As a cell surface ligand, JAG1 plays an important role in the Notch signaling pathway. It has also been reported that JAG1 activates the Notch signaling pathway through interacting with the Notch receptors, such as Notch1, Notch2 and Notch3 [18]. Moreover, the specific function of JAG1/Notch pathway has been found in cervical carcinoma, choriocarcinoma [24] and colorectal cancer [8]. In addition, the carcinogenic effects of Notch2 promoted the metastasis of bladder cancer through EMT [15]. Chen et al. also demonstrated that miR-598 inhibited metastasis in colorectal cancer by suppressing the JAG1/Notch pathway that stimulated EMT [4]. Consistent with previous studies, we found that miR-539 inhibited proliferation and metastasis of WT cells by suppressing JAG1-Notch1/3 expression.
5.Conclusion
In summary, miR-539 was downregulated in WT tissues. And miR-539 inhibited the progression of WT through negatively regulating JAG1-Notch1/3 expression. MiR-539 should be a valuable indicator of the diagnosis and prognosis of WT.
Conflict of interest
The authors declare that they have no competing interests.
References
[1] | N.N. An, J. Shawn, J.P. Peng, M.D. Wu and L.G. Huang, Up-regulation of miR-190b promoted growth, invasion, migration and inhibited apoptosis of Wilms’ tumor cells by repressing the PTEN expression, European Review for Medical and Pharmacological Sciences 22: ((2018) ), 961–969. |
[2] | H. Barak, K. Surendran and S.C. Boyle, The role of Notch signaling in kidney development and disease, Advances in Experimental Medicine and Biology 727: ((2012) ), 99–113. |
[3] | A.E. Brodeur and G.M. Brodeur, Abdominal masses in children: Neuroblastoma, Wilms tumor, and other considerations, Pediatrics in Review 12: ((1991) ), 196–207. |
[4] | J. Chen, H. Zhang, Y. Chen, G. Qiao, W. Jiang, P. Ni, X. Liu and L. Ma, miR-598 inhibits metastasis in colorectal cancer by suppressing JAG1/Notch2 pathway stimulating EMT, Experimental Cell Research 352: ((2017) ), 104–112. |
[5] | L. Chen, W. Zhang, W. Yan, L. Han, K. Zhang, Z. Shi, J. Zhang, Y. Wang, Y. Li, S. Yu, P. Pu, C. Jiang, T. Jiang and C. Kang, The putative tumor suppressor miR-524-5p directly targets Jagged-1 and Hes-1 in glioma, Carcinogenesis 33: ((2012) ), 2276–2282. |
[6] | M. Cui, W. Liu, L. Zhang, F. Guo, Y. Liu, F. Chen, T. Liu, R. Ma and R. Wu, Clinicopathological parameters and prognostic relevance of miR-21 and PTEN expression in Wilms’ tumor, Journal of Pediatric Surgery 52: ((2017) ), 1348–1354. |
[7] | M. Cui, W. Liu, L. Zhang, F. Guo, Y. Liu, F. Chen, T. Liu, R. Ma and R. Wu, Over-expression of miR-21 and lower PTEN levels in Wilms’ tumor with aggressive behavior, The Tohoku Journal of Experimental Medicine 242: ((2017) ), 43–52. |
[8] | Y. Dai, G. Wilson, B. Huang, M. Peng, G. Teng, D. Zhang, R. Zhang, M.P. Ebert, J. Chen, B.C. Wong, K.W. Chan, J. George and L. Qiao, Silencing of Jagged1 inhibits cell growth and invasion in colorectal cancer, Cell Death and Disease 5: ((2014) ), e1170. |
[9] | A.M. Davidoff, Wilms’ tumor, Current Opinion in Pediatrics 21: ((2009) ), 357–364. |
[10] | J.S. Dome, N. Graf, J.I. Geller, C.V. Fernandez, E.A. Mullen, F. Spreafico, M. Van den Heuvel-Eibrink and K. Pritchard-Jones, Advances in Wilms tumor treatment and biology: Progress through international collaboration, Journal of Clinical Oncology: Official Journal of The American Society of Clinical Oncology 33: ((2015) ), 2999–3007. |
[11] | S. Farber, Chemotherapy in the treatment of leukemia and Wilms’ tumor, Jama 198: ((1966) ), 826–836. |
[12] | A. Grovas, A. Fremgen, A. Rauck, F.B. Ruymann, C.L. Hutchinson, D.P. Winchester and H.R. Menck, The National Cancer Data Base report on patterns of childhood cancers in the United States, Cancer 80: ((1997) ), 2321–2332. |
[13] | P.E. Grundy, N.E. Breslow, S. Li, E. Perlman, J.B. Beckwith, M.L. Ritchey, R.C. Shamberger, G.M. Haase, G.J. D’Angio, M. Donaldson, M.J. Coppes, M. Malogolowkin, P. Shearer, P.R. Thomas, R. Macklis, G. Tomlinson, V. Huff and D.M. Green, National Wilms Tumor Study, Loss of heterozygosity for chromosomes 1p and 16q is an adverse prognostic factor in favorable-histology Wilms tumor: A report from the National Wilms Tumor Study Group, Journal of Clinical Oncology: Official Journal of The American Society of Clinical Oncology 23: ((2005) ), 7312–7321. |
[14] | J. Guo, G. Gong and B. Zhang, miR-539 acts as a tumor suppressor by targeting epidermal growth factor receptor in breast cancer, Scientific Reports 8: ((2018) ), 2073. |
[15] | T. Hayashi, K.M. Gust, A.W. Wyatt, A. Goriki, W. Jager, S. Awrey, N. Li, H.Z. Oo, M. Altamirano-Dimas, R. Buttyan, L. Fazli, A. Matsubara and P.C. Black, Not all Notch is created equal: The oncogenic role of Notch2 in bladder cancer and its implications for targeted therapy, Clinical Cancer Research: An Official Journal of The American Association for Cancer Research 22: ((2016) ), 2981–2992. |
[16] | H. Jin and W. Wang, MicroRNA-539 suppresses osteosarcoma cell invasion and migration in vitro and targeting Matrix metallopeptidase-8, International Journal of Clinical and Experimental Pathology 8: ((2015) ), 8075–8082. |
[17] | G.L. Liu, H.J. Yang, B. Liu and T. Liu, Effects of microRNA-19b on the proliferation, apoptosis, and migration of Wilms’ tumor cells via the PTEN/PI3K/AKT signaling pathway, Journal of Cellular Biochemistry 118: ((2017) ), 3424–3434. |
[18] | M.X. Liu, M.K. Siu, S.S. Liu, J.W. Yam, H.Y. Ngan and D.W. Chan, Epigenetic silencing of microRNA-199b-5p is associated with acquired chemoresistance via activation of JAG1-Notch1 signaling in ovarian cancer, Oncotarget 5: ((2014) ), 944–958. |
[19] | Z. Liu, G. Zhang, W. Yu, N. Gao and J. Peng, miR-186 inhibits cell proliferation in multiple myeloma by repressing Jagged1, Biochemical and Biophysical Research Communications 469: ((2016) ), 692–697. |
[20] | J. Lu, G. Song, Q. Tang, J. Yin, C. Zou, Z. Zhao, X. Xie, H. Xu, G. Huang, J. Wang, D.F. Lee, R. Khokha, H. Yang and J. Shen, MiR-26a inhibits stem cell-like phenotype and tumor growth of osteosarcoma by targeting Jagged1, Oncogene 36: ((2017) ), 231–241. |
[21] | L. Miele, Notch signaling, Clinical Cancer Research: An Official Journal of The American Association for Cancer Research 12: ((2006) ), 1074–1079. |
[22] | A. Mirghasemi, A. Taheriazam, S.H. Karbasy, A. Torkaman, M. Shakeri, E. Yahaghi and A. Mokarizadeh, Down-regulation of miR-133a and miR-539 are associated with unfavorable prognosis in patients suffering from osteosarcoma, Cancer Cell International 15: ((2015) ), 86. |
[23] | S. Muthusamy, A.M. DeMartino, L.J. Watson, K.R. Brittian, A. Zafir, S. Dassanayaka, K.U. Hong and S.P. Jones, MicroRNA-539 is up-regulated in failing heart, and suppresses O-GlcNAcase expression, The Journal of Biological Chemistry 289: ((2014) ), 29665–29676. |
[24] | R.T. Pang, C.O. Leung, T.M. Ye, W. Liu, P.C. Chiu, K.K. Lam, K.F. Lee and W.S. Yeung, MicroRNA-34a suppresses invasion through downregulation of Notch1 and Jagged1 in cervical carcinoma and choriocarcinoma cells, Carcinogenesis 31: ((2010) ), 1037–1044. |
[25] | J. Quan, J. Qu and L. Zhou, MicroRNA-539 inhibits glioma cell proliferation and invasion by targeting DIXDC1, Biomedicine and Pharmacotherapy = Biomedecine and Pharmacotherapie 93: ((2017) ), 746–753. |
[26] | M.N. Rivera and D.A. Haber, Wilms’ tumour: Connecting tumorigenesis and organ development in the kidney, Nature Reviews Cancer 5: ((2005) ), 699–712. |
[27] | N. Sethi, X. Dai, C.G. Winter and Y. Kang, Tumor-derived JAGGED1 promotes osteolytic bone metastasis of breast cancer by engaging notch signaling in bone cells, Cancer Cell 19: ((2011) ), 192–205. |
[28] | A.P. South, R.J. Cho and J.C. Aster, The double-edged sword of Notch signaling in cancer, Seminars in Cell and Developmental Biology 23: ((2012) ), 458–464. |
[29] | C.A. Stiller and D.M. Parkin, International variations in the incidence of childhood renal tumours, British Journal of Cancer 62: ((1990) ), 1026–1030. |
[30] | H.F. Wang, Y.Y. Zhang, H.W. Zhuang and M. Xu, MicroRNA-613 attenuates the proliferation, migration and invasion of Wilms’ tumor via targeting FRS2, European Review for Medical and Pharmacological Sciences 21: ((2017) ), 3360–3369. |
[31] | H.J. Xiao, Q. Ji, L. Yang, R.T. Li, C. Zhang and J.M. Hou, In vivo and in vitro effects of microRNA-124 on human gastric cancer by targeting JAG1 through the Notch signaling pathway, Journal of Cellular Biochemistry 119: ((2018) ), 2520–2534. |
[32] | Z.H. Ye and D.W. Gui, miR539 suppresses proliferation and induces apoptosis in renal cell carcinoma by targeting high mobility group A2, Molecular Medicine Reports 17: ((2018) ), 5611–5618. |
[33] | X.B. Zhu, W.J. Lin, C. Lv, L. Wang, Z.X. Huang, S.W. Yang and X. Chen, MicroRNA-539 promotes osteoblast proliferation and differentiation and osteoclast apoptosis through the AXNA-dependent Wnt signaling pathway in osteoporotic rats, Journal of Cellular Biochemistry ((2018) ). |


